Kako ne samo razumjeti značenje strukturnih obilježja stanice, već i glavnu stvar, razumjeti funkcionalne dijelove svojih pojedinačnih komponenti i cijelu stanicu općenito, kombinirati studiju morfologije stanica s glavnim biokemijskim i genetske značajke svog uređaja i rade na proučavanju stanica upravo položaje moderne biologije stanica, potrebno je odmah prisjetiti glavne molekularne biološke obrasce, još jednom se nakratko odnose na sadržaj središnje dogme molekularne biologije.
Stanica kao takva obavlja različite funkcije. Kao što smo već govorili, neki od njih su uglavnom ćelija, neke su posebne, karakteristične za posebne vrste stanica. Glavni radni mehanizmi za obavljanje ovih funkcija su proteini ili njihovi kompleksi s drugim biološkim makromolekulama, kao što su nukleinske kiseline, lipide i polisaharide. Na primjer, poznato je da se transportni procesi u različitim tvarima, počevši s ionima i završetkom makromolekula, određuju se s radom posebnih proteina ili kompleksa lipoproteina koji su dio plazme i drugih staničnih membrana. Gotovo svi procesi sinteze, propadanja, restrukturiranja različitih proteina, nukleinskih kiselina, lipida, ugljikohidrata javljaju se kao rezultat aktivnosti specifične za svaku pojedinačnu reakciju proteina enzima. Sinteza pojedinačnih bioloških monomera, nukleotida, aminokiselina, masnih kiselina, šećera i drugih spojeva također se provode veliki broj specifičnih enzima - proteina. Redukcija koja dovodi do mobilne mobilnosti ili na kretanje tvari i konstrukcija unutar stanica također se provodi posebnim kontraktilnim proteinima. Mnoge stanične reakcije kao odgovor na utjecaj vanjskih čimbenika (virusi, hormoni, stranih proteina, itd.) Počnite s interakcijom tih čimbenika s posebnim receptorskim stanicama.
Proteini su glavne komponente gotovo svih staničnih struktura. Mnoge kemijske reakcije unutar stanice određuju se skup enzima, od kojih svaki dovodi do jedne ili više pojedinačnih reakcija. Struktura svakog pojedinog proteina strogo je specificirana, koja se izražava u specifičnosti njihove primarne strukture - u aminokiselinskoj sekvenci duž polipeptida, proteinskog lanca. Štoviše, specifičnost ove aminokiselinske sekvence se nepogrešivo ponavlja u svim molekulama ovog staničnog proteina.
Takva ispravnost u reproduciranju nedvosmislene sekvence aminokiselina u lancu proteina određena je strukturom DNA dionog dijela gena, što je u konačnici odgovorno za strukturu i sintezu ovog proteina. Ove ideje služe kao glavni postulat molekularne biologije, njegove "dogme". Informacije o budućoj molekuli proteina se prenosi na mjesto njegove sinteze (u ribosomima) posrednikom - informacijske RNA (Irna), čiji je nukleotidni sastav odražava pripravak i sekvencu nukleotida DNA gena. Polipeptidni lanac je izgrađen u ribosomu, sekvenca aminokiselina u kojima se određuje slijedom nukleotida u Irnoj, sekvencu njihovih trojki. Dakle, središnja dogma molekularne biologije naglašava neidislektornost prijenosa informacija: samo iz DNA na protein pomoću međuprodukta veze - Irna (DNA → Irna → Protein). Za neke viruse koji sadrže RNA, lanac prijenosa informacija može ići u skladu s RNA shemom → Irna → protein. Ne mijenja suštinu slučaja, budući da je ovdje određivanja veza također nukleinska kiselina. Inverzni načini određivanja od proteina na nukleinsku kiselinu, do DNA ili RNA su nepoznati.
Kako bi se nastavili na studiju staničnih struktura povezanih sa svim fazama sinteze proteina, moramo ukratko zadržati na glavnim procesima i komponentama koje određuju ovaj fenomen.
Trenutno, na temelju suvremenih ideja o biosintezi proteina, može se dati sljedeća opća glavna shema ovog složenog i višestupanjskog procesa (sl. 16).
Glavni, "tim", uloga u određivanju specifične strukture proteina pripada deoksiribonukleičkoj kiselini - DNA. Molekula DNA je izuzetno duga linearna struktura koja se sastoji od dva međuclaid polimerna lanca. Kompozitni elementi - monomeri - ovi lanci su četiri vrste deoksiribonukleotida, izmjena ili slijeda čiji je jedinstven po lancu i specifičan za svaku DNA molekulu i svaki od njegovih dijelova. Različito dugi dugi dijelovi DNA molekula odgovorni su za sintezu različitih proteina. Tako jedna molekula DNA može odrediti sintezu velikog broja funkcionalno i kemijski različitih stanica stanica. Za sintezu svakog tipa proteina odgovorna je samo određeni dio molekule DNA. Takav dio molekule DNA povezan sa sintezom jednog drugog proteina u stanici često je označen izrazom "cistrona". Trenutno se koncept cijastrona smatra ekvivalentnim konceptom gena. U jedinstvenoj strukturi gena, u određenom sekvencijskom rasporedu njegovih nukleotida duž lanca, zaključeno je sve informacije o strukturi jednog odgovarajućeg proteina.
Iz opće sheme sinteze proteina, vidi se (vidi sliku 16) da je početna stavka iz koje protok informacija za biosintezu proteina u stanici počinje DNA. Stoga je DNA koja sadrži početni zapis informacija koje treba održavati i reproducirati iz ćelije u ćeliju, od generacije do generacije.
Ukratko dodirivanje pitanja o mjestu skladištenja genetskih informacija, tj. Na lokalizaciji DNK u ćeliji možemo reći sljedeće. Odavno je poznato da će, za razliku od svih ostalih komponenti aparata za bijeloziranje, DNA ima posebnu, vrlo ograničenu lokalizaciju: mjesto u stanicama viših (eukariotskih) organizmica će biti jezgra stanica. U nižim (prokariotskim) organizmima koji nemaju ukrašenu jezgru stanica, DNA se također primjećuje od ostatka protoplazme u obliku jedne ili više kompaktnih nukleotidnih formacija. U potpunosti u skladu s tim, eukariotski kernel ili prokariotski nukleodi odavno se smatraju davateljima gena kao jedinstvenog staničnog organoida, koji kontrolira provedbu nasljednih znakova organizama i njihov prijenos na generacije.
Glavno načelo na temu makromolekularne strukture DNA je tzv. Applementarni princip (Sl. 17). Kao što je već spomenuto, molekula DNA se sastoji od dva interclaid lanca. Ti su se lanci međusobno povezani interakcijom njihovih suprotnih nukleotida. U isto vrijeme, prema strukturnim razlozima, postojanje takve strukture samoužnice moguće je samo ako će suprotni nukleotidi oba lanci biti sterički komplementarni, tj. bit će vaša prostorna struktura da se međusobno nadopunjuju. Takvi komplementarni - komplementarni - parovi nukleotida su parni a-T (adenin-timin) i par g. (Gvanin-citozin).
Stoga, prema ovom načelu komplementarnosti, ako imamo određeni slijed četiriju sorti nukleotida u jednom lancu, zatim u drugom lancu, sekvenca nukleotida će se nedvosmisleno odrediti, tako da svaki i prvi lanac će odgovarati Drugi lanac, svaki t prvi lanac - iu drugom lancu, svaki g prvog lanca je C u drugom lancu i svaki c prvi lanac - r u drugom lancu.
Navedeni strukturni princip koji se temelji na strukturi pjene od molekule DNA olakšava razumijevanje točne reprodukcije izvorne strukture, tj. Točna reprodukcija informacija snimljenih u krugovima molekula u obliku određenog slijeda četiriju varijanti nukleotida. Doista, sinteza novih molekula DNA u stanici javlja se samo na temelju već postojećih DNA molekula. U isto vrijeme, dva lanca originalne molekule DNA počinju raspršiti s jednog od krajeva, a na svakom od odvojenih jednokratnih dijelova počinje se skupljati iz drugog lanca prisutnog u mediju slobodnih nukleotida u točno usklađenosti s načelom komplementarnosti. Proces odstupanja između dva lanca originalne molekule DNA nastavlja se i, prema tome, obje lanci nadopunjuju komplementarnim krugovima. Kao rezultat (kao što se može vidjeti na slici 17), umjesto jedne, nastaju dvije molekule DNA, točno identične izvorniku. U svakoj od nastalih "kćer" molekula DNA, jedan lanac je u potpunosti zbog izvornika, a drugi je nerazumno sintetiziran.
Potrebno je naglasiti da je potencijalna sposobnost da se točno reproducira u najmanjoj komplementarnoj komplementarnoj strukturi DNA kao takav, a otkriće toga je definitivno jedno od glavnih postignuća biologije.
Međutim, problem reprodukcije (smanjenje) DNK nije iscrpljen zakonom potencijalne sposobnosti njegove strukture kako bi točno reproducirao njezin nukleotidni slijed. Činjenica je da sama DNA uopće nije samo-reproducijska molekula. Za provedbu procesa sinteze - reprodukcija DNA prema gore opisanoj shemi - aktivnost posebnog enzimskog kompleksa, koji je potreban naziv DNA polimeraze. To je taj enzim da je molekula DNA dosljedno govori iz jednog kraja u drugi proces odstupanja dvaju lanca s istovremenom polimerizacijom na njima slobodne nukleotide u skladu s komplementarnim načelom. Dakle, DNA, poput matrice, samo postavlja redoslijed nukleotida u sintetizirajućim lancima, a sam proces vodi protein. Rad enzima tijekom smanjenja DNA danas je jedan od najzanimljivijih problema. Vjerojatno je DNA polimeraza kao što bi trebalo aktivno puzati duž dvadeset molekule DNA s jednog kraja na drugu, ostavljajući iza split smanjenog "repa" iza. Fizički principi ovog rada ovog proteina još nisu jasni.
Međutim, DNA i njegovi pojedinačni funkcionalni dijelovi koji nose informacije o strukturi proteina, sami izravan sudjelovanje u procesu stvaranja proteinskih molekula se ne prihvaćaju. Prva faza na putu do provedbe tih informacija zabilježenih u DNA lancima je tzv proces transkripcije ili "prepisivanje". U tom procesu, na istom DNK krugu, kao i na matrici, dolazi do sinteze kemijski povezane polimerne kiseline (RNA). Molekula RNA je jedan lanac, čiji su monomeri četiri vrste ribonukleotida, koji se smatraju malom modifikacijom četiriju sorti DNA deoksiribonukleotida. Slijed četiriju sorti ribonukleotida u dobivenom RNA krugu točno ponavlja sekvencu odgovarajućih deoksiribonukleotida jednog od dva DNA krugova. Na taj način, nukleotidna sekvenca gena se kopira kao RNA molekule, tj. Informacije zabilježene u strukturi ovog gena u potpunosti su prepiše na RNA. Veliki, teoretski neograničen broj takvih "kopija" - RNA molekule mogu se ukloniti iz svakog gena. Ove molekule, prepisane u mnogim kopijama "kopija" gena i, da se nose iste informacije kao geni, razlikuju se kroz kavez. Oni su već izravno uključeni u stanice stanica i prihvate "osobno" sudjelovanje u procesu stvaranja proteinskih molekula. Drugim riječima, oni nose informacije s mjesta gdje se pohranjuje u njegovoj provedbi. Sukladno tome, ove RNA označava kao informativnu (IRNA) ili matricu (mRNA).
Utvrđeno je da se lanac IRNN sintetizira, izravno koristeći odgovarajući DNA sekciju kao matricu. Sintetizirani mRNA krug u isto vrijeme točno kopira jedan od dva DNA krugova po njegovoj nukleotidnom sekvenci (uzimajući da uracil (y) u RNA odgovara njegovom timinskom derivatu (t) u DNA). To se događa na temelju istog strukturnog načela komplementarnosti, koji određuje smanjenje DNA (Sl. 18). Pokazalo se da kada se pojavi sinteza mRNA na DNA u stanici, tada se koristi samo jedan lanac DNA kao matrica za formiranje mRNA kruga. Zatim, svaki g ovog DNK kruga će odgovarati C u lancu RNA u konstrukciji, svaki C lanca DNA kruga u RNA krugu, svaki t lanac - A u RNA krugu i svaki DNA krug u RNA krugu. Kao rezultat toga, dobiveni RNA lanac će biti strogo komplementaran lancem matriksa DNA i stoga je identičan nukleotidnom sekvenci (uzimajući t \u003d y) drugi lanac DNA. Dakle, "prepisivanje" informacija iz DNA na RNA javlja, tj. transkripcija. Kombinacije "prepisane" nukleotida RNA lanca već su izravno određene rasporedom odgovarajućih, kodiranih aminokiselina u lancu proteina.
Ovdje, kao iu razmatranju smanjene DNK, kao jedan od najznačajnijih trenutaka procesa transkripcije, potrebno je ukazati na njegovu enzimsku prirodu. DNA, koja je matrica u ovom procesu, određuje cijeli nukleotidni položaj u sintetizirajućem lancu mRNA, sve specifičnosti RNA formira, ali sam proces se provodi posebni protein - enzim. Ovaj enzim se naziva RNA polimeraza. Njegova molekula ima složenu organizaciju koja mu omogućuje da se aktivno kreće duž molekule DNA, u isto vrijeme sintetizirajući lanac RNA komplementarne jednom od DNA lanca. DNA molekula, koja služi kao matrica, ne troši i ne mijenja, ostaje u istom obliku i uvijek je spreman za takav prepisivanje od njega neograničen broj "kopija" - mRNA. Protok tih mRNA iz DNA do ribosoma je protok informacija koje osiguravaju programiranje stanica stanice stanice, cijelu cjelokupnu cjelokupnost svojih ribosoma.
Prema tome, razmatrani dio sheme opisuje protok informacija koje proizlaze iz DNA u obliku mRNA molekula na intracelularne čestice sintetiziraju proteine. Sada se okrećemo struji druge vrste - tok materijala iz kojeg bi se trebao stvoriti protein. Elementarne jedinice - monomeri - Molekula proteina su aminokiseline, koje su numerirane oko 20. Za stvaranje proteinske molekule, slobodne aminokiseline prisutne u stanici moraju biti uključeni u odgovarajuću tok koji ulazi u česticu bijelooksiatiranja, a već postoje u lancu s određenim jedinstvenim načinom., diktirane informacije RNA. Takvo sudjelovanje aminokiselina - građevinski materijal za stvaranje proteina se provodi prianjanjem slobodnih aminokiselina u posebne RNA molekule relativno male. Ove rne koje služe za prilogu slobodnih aminokiselina s njima bez informacijske, nose drugačije - adapter - funkciju, što znači dalje biti vidljivo. Aminokiseline se spoje jedan od krajeva malih lanaca transfera RNA (TRNA), jednu aminokiselinu po molekuli RNA. Za svaku takvu aminokiselinu u stanici postoje njegove specifične adapterne RNA molekule koje pričvršćuju samo ove aminokiseline. U takvom montiranom obliku aminokiselina na RNA i uđe u čestice bijelooksiatiranja.
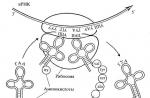
Središnji trenutak procesa biosinteze proteina je fuzija ova dva intracelularna toka - protok informacija i protok materijala - u stanicama stanica. Te se čestice nazivaju ribosomi. Ribosomi su ultramikroskopski biokemijski "strojevi" molekularnih veličina, gdje se specifični proteini prikupljaju iz dolaznih aminokiselinskih ostataka, prema planu zaključenoj u informaciji RNA. Iako na Sl. 19 prikazuje samo jednu česticu, svaka ćelija će zadržati tisuće rebara. Količina ribosoma određuje ukupni intenzitet sinteze proteina u stanici. Promjer jedne ribosomalne čestice je oko 20 nm. Kemijskom prirodom ribosoma - ribonukleoteida: sastoji se od posebne ribosomalne RNA (to je treća RNA klasa koja nam je poznavala uz informacije i adapter RNA) i molekule strukturnog ribosomalnog proteina. Zajedno, ova kombinacija nekoliko desetaka makromolekula formira idealno organiziranog i pouzdanog "automobila", koji ima nekretninu za čitanje informacija sklopljenih u lancu MRNA, te da je shvati u obliku gotove proteinske molekule određene strukture. Budući da je stvorenje procesa je li linearni raspored 20 različitih aminokiselina u lancu proteina nedvosmisleno određen položajem četiri različita nukleotida u lancu kemijski potpuno drugačije polimerne kiseline (mRNA), ovaj proces koji se pojavljuje u Ribosome je napravljen za označavanje termina "emitiranja", ili "prijevod" - prijevod iz četveroslovnog abecede lanca nukleinskih kiselina u dvadeset i zabilježena abeceda proteina (polipeptidnih) lanca. Kao što se može vidjeti, sva tri poznata RNA klase su uključene u proces emitiranja: informacija RNA, koja je objekt emitiranja; Ribosomalna RNA, koja igra ulogu organizatora anti-industrijskog ribonukleotopoodnih čestica - ribosoma; I adapter RNA obavlja funkciju prevoditelja.
|
Sl. 19. Shema funkcioniranja ribosoma |

Proces sinteze proteina započinje formiranjem spojeva aminokiselina s adapterom RNA molekulama ili Trnom. U isto vrijeme, energija "aktivacija" aminokiselina se najprije dogodila zbog njegove enzimske reakcije s adenozinom trifhosfat molekulom (ATP), a zatim je "aktivirana" aminokiselina povezana s kraja relativno kratkotrajne TRNA. Lanac, povećanje kemijske energije aktivirane aminokiseline je intenzitet u obliku energije kemijske veze između aminokiseline i Trne.
U isto vrijeme, drugi zadatak je riješen. Činjenica je da se reakcija između aminokiseline i molekule TRNA provodi enzim označen kao sintetaza visok aminacil. Za svaku od 20 aminokiselina postoje posebni enzimi koji reagiraju uz sudjelovanje samo ove aminokiseline. Prema tome, postoji najmanje 20 enzima (aminoacil-visok sintetaza), od kojih je svaki specifičan za jednu specifičnu aminokiselinu. Svaki od ovih enzima može imati reakciju ne s bilo kojom TRNA molekulom, već samo s onima koji nose strogo definiranu kombinaciju nukleotida u njihovom lancu. Dakle, zbog postojanja skupa takvih specifičnih enzima koji se razlikuju, s jedne strane, priroda aminokiseline i, na drugoj - nukleotidna sekvenca Trvine, svaka od 20 aminokiselina se ispostavlja kao " dodijeljen "samo na određenu TRNA s određenom karakterističnom kombinacijom nukleotida.
Neki trenuci procesa biosinteze proteina, koliko i danas predstavljamo, daju se na Sl. 19. Ovdje, prije svega, može se vidjeti da je molekula informacija RNA spojena na ribosomu ili, kako kažu, ribosomu je "programiran" informacija RNA. U danom trenutku, izravno u samom ribosomu samo je relativno kratak segment lanca mRNA. Ali to je taj segment uz sudjelovanje ribosoma može komunicirati s adapterom RNA molekule. I opet je glavna uloga odigrana načelom komplementacije.
Ovo je objašnjenje mehanizma zašto ta tript lanca mRNA odgovara strogo definiranoj aminokiselini. Potrebna srednja veza ili adapter, s "prepoznavanjem" svake aminokiseline njegovog trojke na mRNA je adapter RNA (TRNA).
Na sl. 19 Može se vidjeti da u ribosomu, uz molekulu TRNA s višom aminokiselinom, postoji još jedna Trna molekula. No, za razliku od gore spomenute molekule TRNA, ova molekula TRNA je pričvršćena na kraj do kraja sinteze lanca proteina (polipeptida). Ova odredba odražava dinamiku događaja koji se pojavljuju u ribosomu tijekom sinteze proteinskih molekula. Ova dinamika može se zamisliti kako slijedi. Počnimo s određenim srednjim zakretnim momentom koji se odražava na sl. 19 i karakterizira prisutnost već početka da se gradi proteinski lanac pričvršćen na njezin TRNA i upravo ušao u ribosoma i povezan s trojkom nove molekule TRNA s odgovarajućom aminokiselinom. Očigledno, čin vezivanja TRNA molekule na ribosomu koji se nalazi ribosomski triplet dovodi do takve međusobne orijentacije i bliski kontakt između aminokiselinskog ostatka i proteinskog kruga u konstrukciji, koji se javlja kovalentna veza između njih. Komunikacija nastaje na takav način da je kraj proteinskog lanca u izgradnji (na slici 19 pričvršćen na TRNA) se prenosi iz ovog TRNA na aminokiselinski ostatak aminoacilnog trgovanja. Kao rezultat "desne" TRNA, igranje uloge "donatora", bit će slobodan, a protein lanac - prenesen u "akceptor", tj. Na "lijevo" (primljeno) aminocil-TRNA. Kao rezultat toga, protein lanac će biti izdužen po aminokiselini i pričvršćen na "lijevo" trt. Slijedeći to, "lijevo" TRNA se prenosi zajedno s trojkom povezanim s nukleotidom mRNA udesno, a zatim će odavde biti zamjen bivše "donator" TRNA molekula i ostavlja ribosome. Na njezinu mjestu, pojavljuje se novi TRNA s proteinskim lancem u konstrukciji produžen na jedan aminokiselinski ostatak, a mRNA lanac će se promicati u odnosu na ribosomu na jednom tripletu desno. Kao rezultat promocije lanca mRNA, jedan triplet u desnoj strani u ribosomu će se pojaviti sljedeći prazan trotić (UOU), a odgovarajući TRNA s aminokiselinom (fenilalanil-TRNA) odmah će biti dodan u skladu s komplementarno načelo. To će ponovno uzrokovati stvaranje kovalentne (peptidne) veze između lanca proteina u konstrukciji i ostatku fenilalanina i slijedi promociju lanca mRNA do jednog tripleta s desne strane sa svim posljedicama koje proizlaze odavde, itd. Na taj se način provode dosljedno, trojku za trojku, povlačeći lanac informacija RNA kroz ribosomu, kao posljedica toga što je lanca IRNN-a u cijelosti "pročitao" od početka do kraja. U isto vrijeme, postoji dosljedna aminokiselina za aminokiselinu, lanca proteina konjugu. Prema tome, u ribosomu, jedan za drugom dolazi molekule Trvine s aminokiselinama, a molekule TRNA izazivaju bez aminokiselina. Popravak u otopini izvan ribosoma, slobodne molekule TRNA ponovno su povezane s aminokiselinama i ponovno ih nose u ribosomu, čime se ciklički odnosi bez razaranja i promjena.
Glavni postulat molekularne biologije
Postoje tri molekularna procesa biologije.
Stečen
Može se promatrati ovaj češći oblik Orotatacidurije:
· S bilo kakvim nedostatkom enzimi sinteze ureeosim karbamoil fosfatne sintetaze. U isto vrijeme, iz njih proizlazi mitohondrijski karbamoil fosfat (u normi koja se koristi za stvaranje ureje) i koristi se za prekomjerne sinteze kiseline. Bolest je obično popraćena bygipmermonMiamia,
· U liječenju alopurinol gihta, koji se može pretvoriti u oksiparinomonukleotid, koji je inhibitor orotatdekarboksilaze, koji opet dovodi do nakupljanja Orotata.
Glavna figura matrične biosinteze su nukleinska kiselina RNA i DNA. Oni su polimerne molekule koje uključuju dušične baze od pet vrsta, pentoze od dva tipa i ostaci fosforne kiseline. Baze dušika u nukleinskim kiselinama mogu biti purina ( adenin,gvajka) i pirimidin ( citozin, uracil (samo u RNA) timin (samo u DNK)). Ovisno o strukturi ugljikohidrata, označite ribonukleinske kiseline - sadrži Ribosa (RNA) i deoksiribonukleinske kiseline - sadržavati dezoksiribozu (DNA).
Pojam " matrična biosinteza"To podrazumijeva sposobnost ćelije da sintetizira polimerne molekule, kao što je nukleinske kiseline i proteini, na temelju predloška - matrica, Time se osigurava točan prijenos najsloženije strukture od već postojećih molekula do novoizmjenjivih.
U velikoj većini slučajeva, prijenos nasljednih informacija iz majčinske ćelije na podružnicu provodi se pomoću DNK ( replikacija). Koristiti genetske informacije, sama stanica zahtijeva RNA formirana na DNA matrici ( transkripcija). Nadalje, RNA je izravno uključena u sve faze sinteze proteinskih molekula ( emitiranje), osiguravajući strukturu i aktivnost ćelije.
Na gore navedeno središnja molekularna biologijaPrema kojem se prijenos genetskih informacija provodi samo na nukleinskoj kiselini (DNA i RNA). Primatelj informacija može biti druga nukleinska kiselina (DNA ili RNA) i proteina.
Hibridizacija je već široko korištena
Ako se zagrijete RNA otopinu iznad temperature od 90 ° C ili pomaknute pH u oštroalkalne ili oštro, vodikove veze između DNA niti su uništene, a dvostruka spirala je slomljena. Nastaje denaturacija DNAili, na drugačiji način, topljenje, Ako izbrišete agresivni faktor, onda se događa renaturaili žarenje, Tijekom žarenja, DNK niti "pronalaze" komplementarne površine jedna od druge i, na kraju se ponovno pojavljuju u dvostruku spiruju.
Ako u jednoj "cijevi" taljenje i žarenjem DNA smjese, na primjer, čovjeki mišNeki dijelovi miša DNK krugova će se ponovno ujediniti s komplementarnim područjima humanih DNA lanca s obrazovanjem hibrida, Broj takvih web-lokacija ovisi o stupnju srodstva vrsta. Bliže stavove između sebe, to je veća komplementarnost DNA niti. Ovaj fenomen se zove hibridizacija DNA DNA.
Ako je RNA prisutna u otopini, onda možete implementirati hibridizacija DNA RNA, Takva hibridizacija pomaže uspostaviti blizinu određenih DNA sekvenci s bilo kojom RNA.
Hibridizacija DNA DNA i DNA RNA se koristi kao učinkovit alat U molekularnoj genetici, forenzička medicina, antropologija uspostavljanja genetskog srodstva između vrsta.
Stanica kao takva posjeduje ogroman broj različitih funkcija, kao što smo već govorili, neki od njih su općenito stanica, neki su posebni, karakteristični za posebne vrste stanica. Glavni radni mehanizmi za obavljanje ovih funkcija su proteini ili njihovi kompleksi s drugim biološkim makromolekulama, kao što su nukleinske kiseline, lipide i polisaharide. Dakle, zna se da su procesi prijevoza u različitim tvarima, počevši s ionima, završavajući s makromolekulama, određuju se djelovanjem posebnih proteina ili kompleksa lipoproteina u sastavu plazme i drugih staničnih membrana. Praktički svi procesi sinteze, propadanja, restrukturiranja različitih proteina, nukleinskih kiselina, lipida, ugljikohidrata nastaje kao rezultat djelovanja specifične za svaku pojedinačnu reakciju proteina enzima. Sinteza pojedinačnih bioloških monomera, nukleotida, aminokiselina, masnih kiselina, šećera itd. Također se provode veliki broj specifičnih enzima - proteina. Redukcija koja dovodi do mobilne mobilnosti ili na kretanje tvari i konstrukcija unutar stanica također se provodi posebnim kontraktilnim proteinima. Mnoge stanične reakcije kao odgovor na utjecaj vanjskih čimbenika (virusa, hormona, vanzemaljskih proteina, itd.) Počinje interakcijom tih čimbenika s posebnim staničnim receptorima.
Proteini su glavne komponente gotovo svih staničnih struktura. Mnoge kemijske reakcije unutar stanice određuju se skup enzima, od kojih svaki dovodi do jedne ili više pojedinačnih reakcija. Struktura svakog pojedinog proteina strogo je specificirana, koja se izražava u specifičnosti njihove primarne strukture - u aminokiselinskoj sekvenci duž polipeptida, proteinskog lanca. Štoviše, specifičnost ove aminokiselinske sekvence se nepogrešivo ponavlja u svim molekulama ovog staničnog proteina.
Takva ispravnost u reproduciranju nedvosmislene sekvence aminokiselina u lancu proteina određena je strukturom DNA dionog dijela gena, što je u konačnici odgovorno za strukturu i sintezu ovog proteina. Ove ideje služe kao glavni postulat molekularne biologije, njegove "dogme". Informacije o budućoj molekuli proteina se prenosi na mjesto njegove sinteze (u ribosomima) posrednikom - informacijske RNA (Irna), čiji je nukleotidni sastav odražava pripravak i sekvencu nukleotida DNA gena. Polipeptidni lanac je izgrađen u ribosomu, sekvenca aminokiselina u kojima se određuje slijedom nukleotida u Irnoj, sekvencu njihovih trojki. Dakle, središnja dogma molekularne biologije naglašava neidisleksnost prijenosa informacija: samo iz DNA do proteina, koristeći intermedijer, IRNA (DNA ® IRNK ® protein). Za neke viruse koji sadrže RNA, promet za prijenos informacija može ići u skladu s RNA shemom - IRNK - proteinom. Ne mijenja suštinu slučaja, budući da je ovdje određivanja veza također nukleinska kiselina. Inverzni načini određivanja od proteina na nukleinsku kiselinu, do DNA ili RNA su nepoznati.
Kako bi se nastavili na studiju staničnih struktura povezanih sa svim fazama sinteze proteina, moramo ukratko zadržati na glavnim procesima i komponentama koje određuju ovaj fenomen.
Trenutno, na temelju suvremenih ideja o biosintezi proteina, može se dati sljedeća opća glavna shema ovog složenog i višestupanjskog procesa (sl. 16).
Glavni, "tim", uloga u određivanju specifične strukture proteina pripada deoksiribonukleičkoj kiselini - DNA. Molekula DNA je izuzetno duga linearna struktura koja se sastoji od dva međuclaid polimerna lanca. Kompozitni elementi - monomeri - ovi lanci su četiri vrste deoksiribonukleotida, alternation ili sekvenca od kojih je duž lanca jedinstven i specifičan za svaku DNA molekulu i svaki od njegovih dijelova. Različito dugi dugi dijelovi DNA molekula odgovorni su za sintezu različitih proteina. Tako jedna molekula DNA može odrediti sintezu velikog broja funkcionalno i kemijski različitih stanica stanica. Za sintezu svakog tipa proteina odgovorna je samo određeni dio molekule DNA. Takav dio molekule DNA povezane sa sintezom jednog proteina u stanici često je označen pojmom "cystron". Trenutno se koncept cijastrona smatra ekvivalentnim konceptom gena. U jedinstvenoj strukturi gena, u određenom sekvencijskom rasporedu njegovih nukleotida duž lanca, zaključeno je sve informacije o strukturi jednog odgovarajućeg proteina.
Iz opće sheme sinteze proteina, vidi se (vidi sliku 16) da je početna stavka iz koje protok informacija za biosintezu proteina u stanici počinje DNA. Stoga je DNA koja sadrži početni zapis informacija koje treba održavati i reproducirati iz ćelije u ćeliju, od generacije do generacije.
Ukratko dodirivanje pitanja o mjestu skladištenja genetskih informacija, tj. Na lokalizaciji DNK u ćeliji možemo reći sljedeće. Odavno je poznato da će, za razliku od svih ostalih komponenti aparata za bijeloziranje, DNA ima posebnu, vrlo ograničenu lokalizaciju: mjesto u stanicama viših (eukariotskih) organizmica će biti jezgra stanica. U nižim (prokariotskim) organizmima koji nemaju ukrašenu jezgru stanica, DNA se također primjećuje od ostatka protoplazme u obliku jedne ili više kompaktnih nukleotidnih formacija. U potpunosti u skladu s tim, eukariotski kernel ili prokariotski nukleodi odavno se smatraju davateljima gena kao jedinstvenog staničnog organoida, koji kontrolira provedbu nasljednih znakova organizama i njihov prijenos na generacije.
Osnovno načelo na temu makromolekularne strukture DNA je takozvani komplementarni princip (Sl. 17). Kao što je već spomenuto, molekula DNA se sastoji od dva interclaid lanca. Ti su se lanci međusobno povezani interakcijom njihovih suprotnih nukleotida. U isto vrijeme, prema strukturnim razlozima, postojanje takve strukture samoužnice moguće je samo ako će suprotni nukleotidi oba lanci biti sterički komplementarni, tj. bit će vaša prostorna struktura da se međusobno nadopunjuju. Takvi komplementarni - komplementarni - parovi nukleotida su parni a-T (adenin-timin) i par g. (Gvanin-citozin).
Stoga, prema ovom načelu komplementarnosti, ako imamo određeni slijed četiriju sorti nukleotida u jednom lancu, zatim u drugom lancu, sekvenca nukleotida će se nedvosmisleno odrediti, tako da svaki i prvi lanac će odgovarati Drugi lanac, svaki t prvi lanac - iu drugom lancu, svaki g prvog lanca je C u drugom lancu i svaki c prvi lanac - r u drugom lancu.
Može se vidjeti da je navedeno strukturno načelo na kojoj se temelji struktura pjene od molekule DNA olakšava razumijevanje točne reprodukcije izvorne strukture, tj. Točna reprodukcija informacija zabilježenih u krugovima molekula u obliku određenog slijeda od 4 sorte nukleotida. Doista, sinteza novih molekula DNA u stanici javlja se samo na temelju već postojećih DNA molekula. U isto vrijeme, dva lanca originalne molekule DNA počinju raspršiti s jednog od krajeva, a na svakom od odvojenih jednokratnih dijelova počinje se skupljati iz drugog lanca prisutnog u mediju slobodnih nukleotida u točno usklađenosti s načelom komplementarnosti. Proces odstupanja između dva lanca originalne molekule DNA nastavlja se i, prema tome, obje lanci nadopunjuju komplementarnim krugovima. Kao rezultat toga, kao što se vidi na dijagramu, dvije molekule DNA pojavljuju umjesto jedne, točno identične izvorniku. U svakom od nastalih "podružnica" molekula DNA, jedan lanac, kao što se vidi, u potpunosti je zbog izvornika, a drugi je aow sintetiziran.
Glavna stvar je da je potrebno ponovno naglasiti, to je da se potencijalna sposobnost da se točno reproducira u najmanjoj komplementarnoj strukturi DNK, a otkriće je to svakako jedno od glavnih postignuća biologije.
Međutim, problem reprodukcije (smanjenje) DNK nije iscrpljen zakonom potencijalne sposobnosti njegove strukture kako bi točno reproducirao njezin nukleotidni slijed. Činjenica je da sama DNA uopće nije samo-reproducijska molekula. Provesti proces sinteze - reprodukcija DNA prema gore opisanoj shemi, potrebna je aktivnost posebnog enzimskog kompleksa koji se naziva DNA polimeraza. Očigledno, to je taj enzim da je molekula DNA dosljedno govori iz jednog kraja u drugi proces odstupanja dvaju krugova s \u200b\u200bistovremenom polimerizacijom na njima slobodne nukleotide komplementarnim principom. Dakle, DNA, poput matrice, samo postavlja redoslijed nukleotida u sintetizirajućim lancima, a sam proces vodi protein. Rad enzima tijekom smanjenja DNA danas je jedan od najzanimljivijih problema. Očigledno, čini se da je DNA polimeraza aktivno puzala duž najgnutnije molekule DNA od jednog kraja na drugu, ostavljajući iza split smanjenog "repa". Fizički principi ovog rada ovog proteina još nisu jasni.
Međutim, DNA i njegovi pojedinačni funkcionalni dijelovi koji nose informacije o strukturi proteina, sami izravan sudjelovanje u procesu stvaranja proteinskih molekula se ne prihvaćaju. Prva faza na putu do provedbe tih informacija zabilježenih u DNA lancima je tzv proces transkripcije ili "prepisivanje". U tom procesu, DNK lanci, kao i na matrici, postoji sinteza kemijski povezanog polimera - ribonukleivna kiselina (RNA). Molekula RNA je jedan lanac, čiji su monomeri četiri vrste ribonukleotida, koji se smatraju malom modifikacijom četiriju sorti DNA deoksiribonukleotida. Slijed četiriju sorti ribonukleotida u dobivenom RNA krugu točno ponavlja sekvencu odgovarajućih deoksiribonukleotida jednog od dva DNA krugova. Na taj način, nukleotidna sekvenca gena se kopira kao RNA molekule, tj. Informacije zabilježene u strukturi ovog gena u potpunosti su prepiše na RNA. Veliki, teoretski neograničen broj takvih "kopija" - RNA molekule mogu se ukloniti iz svakog gena. Ove molekule, prepisane u mnogim kopijama "kopija" gena i stoga se nose iste informacije kao geni, razlikuju se kroz ćeliju. Oni su već izravno uključeni u stanice stanica i prihvate "osobno" sudjelovanje u procesu stvaranja proteinskih molekula. Drugim riječima, oni nose informacije s mjesta gdje se pohranjuje u njegovoj provedbi. U skladu s tim, ove RNA označava informativne ili matrice RNA, skraćene mRNA (ili IRNA).
Utvrđeno je da je informiranje RNA krug sintetiziran, izravno koristeći odgovarajući DNA dio kao matricu. Sintetizirani mRNA krug u isto vrijeme točno kopira jedan od dva DNA krugova po njegovoj nukleotidnom sekvenci (uzimajući da uracil (y) u RNA odgovara njegovom timinskom derivatu (t) u DNA). To se događa na temelju istog strukturnog načela komplementarnosti, koji određuje smanjenje DNA (Sl. 18). Pokazalo se da kada se pojavi sinteza mRNA na DNA u stanici, tada se koristi samo jedan lanac DNA kao matrica za formiranje mRNA kruga. Zatim, svaki g ovog DNK kruga će odgovarati C u lancu RNA u konstrukciji, svaki C lanca DNA kruga u RNA krugu, svaki t lanac - A u RNA krugu i svaki DNA krug u RNA krugu. Kao rezultat toga, rezultirajući lanac RNA će biti strogo komplementaran lancu DNA matriksa i stoga je identičan na slijedu nukleotida (uzimajući t \u003d y) drugi lanac DNA. Dakle, "prepisivanje" informacija iz DNA na RNA javlja, tj. transkripcija. Kombinacije "prepisane" nukleotida RNA lanca već su izravno određene rasporedom odgovarajućih, kodiranih aminokiselina u lancu proteina.
Ovdje, kao iu razmatranju smanjene DNK, kao jedan od najznačajnijih trenutaka procesa transkripcije, potrebno je ukazati na njegovu enzimsku prirodu. DNA, koja je matrica u ovom procesu, određuje cijeli nukleotidni položaj u sintetizirajućem lancu mRNA, sve specifičnosti RNA formira, ali sam proces se provodi posebni protein - enzim. Ovaj enzim se naziva RNA polimeraza. Njegova molekula ima složenu organizaciju koja mu omogućuje da se aktivno kreće duž molekule DNA, u isto vrijeme sintetizirajući lanac RNA komplementarne jednom od DNA lanca. Molekula DNA, koja služi kao matrica, ne troši i ne mijenja, preostaje u istom obliku i uvijek je spreman za takav prepisivanje od njega neograničen broj "kopija" - mRNA. Protok tih mRNA iz DNA do ribosoma je protok informacija koje osiguravaju programiranje stanica stanice stanice, cijelu cjelokupnu cjelokupnost svojih ribosoma.
Prema tome, razmatrani dio sheme opisuje protok informacija koje proizlaze iz DNA u obliku mRNA molekula na intracelularne čestice sintetiziraju proteine. Sada se okrećemo struji druge vrste - tok materijala iz kojeg bi se trebao stvoriti protein. Osnovne jedinice - monomeri - Molekula proteina su aminokiseline koje imaju 20 različitih sorti. Da bi se stvorila (sintetiziranje) molekula proteina, slobodne aminokiseline prisutne u stanici trebaju biti uključeni u odgovarajuću tok koji ulazi u česticu bijelooksiatiranja, a već su postavljeni u lancu s određenim jedinstvenim načinom diktirane informacijama. Ovo sudjelovanje aminokiselina - građevinski materijal za stvaranje proteina provodi se dodavanjem slobodnih aminokiselina do posebnih RNA molekula relativno malih. Ove RNA koje služe da im se pridruže slobodne aminokiseline neće biti informativne i nose drugačiji adapter - funkcija, što znači da će biti vidljivo. Aminokiseline se spoje jedan od krajeva malih lanaca transfera RNA (TRNA), jednu aminokiselinu po molekuli RNA.
Za svaku raznolikost aminokiselina u ćeliji postoje njezini specifični, koji pridaju samo ovu raznolikost aminokiselina adaptera RNA molekule. U takvom obliku koji se posjeti na RNA, aminokiselinama i ulazi u bijelo oči.
Središnji trenutak procesa biosinteze proteina je fuzija ova dva intracelularna toka - protok informacija i protok materijala - u stanicama stanica. Te se čestice nazivaju ribosomi. Ribosomi su ultramikroskopski biokemijski "strojevi" molekularnih veličina, gdje se specifični proteini prikupljaju iz dolaznih aminokiselinskih ostataka, prema planu zaključenoj u informaciji RNA. Iako u ovoj shemi (sl. 19) prikazuje samo jednu česticu, svaka ćelija će zadržati tisuće rebara. Količina ribosoma određuje ukupni intenzitet sinteze proteina u stanici. Promjer jedne ribosomalne čestice je oko 20 nm. Kemijskom prirodom ribosoma - ribonukleoteida: sastoji se od posebne ribosomalne RNA (to je treća RNA klasa koja nam je poznavala uz informacije i adapter RNA) i molekule strukturnog ribosomalnog proteina. Zajedno, ova kombinacija nekoliko desetaka makromolekula formira idealno organiziranog i pouzdanog "automobila", koji ima nekretninu za čitanje informacija sklopljenih u lancu MRNA, te da je shvati u obliku gotove proteinske molekule određene strukture. Budući da je stvorenje procesa je da je linearni raspored aminokiselina u proteinskom lancu jedinstveno određen položajem četiriju sorti nukleotida u kemijski potpuno različitom polimeru polimera (mRNA), tada se taj proces pojavljuje u Ribosome je napravljen za označavanje termina "emitiranja", ili "prijevod" - prijevod od 4-slova abeceda lanca nukleinskih kiselina od 20 slova abeceda proteina (polipeptida) lanca. Kao što se može vidjeti, sva tri poznata RNA klase su uključene u proces emitiranja: informativna RNA, koja je prevodni objekt, ribosomalna RNA, koja igra ulogu organizatora anti-martr-marvotonukleotopootske čestice - ribosomu i adapter RNA obavljanje funkcije prevoditelja.
Proces sinteze proteina započinje formiranjem spojeva aminokiselina s adapterom RNA molekulama ili Trnom. U isto vrijeme, energija "aktivacija" aminokiselina se najprije dogodila zbog njegove enzimske reakcije s adenozinom trifhosfat molekulom (ATP), a zatim je "aktivirana" aminokiselina povezana s kraja relativno kratkotrajne TRNA. Lanac, povećanje kemijske energije aktivirane aminokiseline je intenzitet u obliku energije kemijske veze između aminokiseline i Trne.
Ali u isto vrijeme, drugi zadatak je riješen. Činjenica je da se reakcija između aminokiseline i molekule TRNA provodi enzim označen kao sintetaza visok aminacil. Za svaku od 20 sorti aminokiselina postoje posebni enzimi koji reagiraju uz sudjelovanje samo ove aminokiseline. Prema tome, postoji najmanje 20 enzima (aminoacil-Trnk sintetaza), od kojih je svaki specifičan za jednu raznolikost aminokiselina. Svaki od ovih enzima može imati reakciju ne s bilo kojom TRNA molekulom, već samo s onima koji nose strogo definiranu kombinaciju nukleotida u njihovom lancu. Dakle, zbog postojanja skupa takvih specifičnih enzima koji se razlikuju, s jedne strane, priroda aminokiseline i, na drugoj - nukleotidnu sekvencu Trvine, svaka od 20 sorti aminokiselina biti "dodijeljen" samo na određenu TRNA s određenom karakterističnom kombinacijom nukleotida.
Neki trenuci procesa biosinteze proteina, koliko i danas predstavljamo, daju se na Sl. devetnaest.
Ovdje, prije svega, može se vidjeti da je informacijska molekula RNA spojena na ribosomu ili, kao što kažu, ribosomu "programira" Informacije RNA. U bilo kojem trenutku, izravno u samom ribosomu samo je relativno kratki segment lanca mRNA. Ali to je taj segment uz sudjelovanje ribosoma može komunicirati s adapterom RNA molekule. I opet se glavna uloga odigra dvostruko navedeno načelo komplementacije.
Ovo je objašnjenje mehanizma zašto ta tript lanca mRNA odgovara strogo definiranoj aminokiselini. Može se vidjeti da je potrebna srednja veza ili adapter, s "prepoznavanjem" svake amino kiseline njezina tripleta na mRNK-u adapter RNA (TRNA).
Nadalje u dijagramu (vidi sl. 19) Može se vidjeti da u ribosomu, uz gore navedene trnalne molekule s višom aminokiselinom, postoji još jedna trna molekula. No, za razliku od gore spomenute molekule TRNA, ova molekula Trna pričvršćena je na kraj proteina (polipeptidnog) lanca u procesu sinteze. Ova odredba odražava dinamiku događaja koji se pojavljuju u ribosomu tijekom sinteze proteinskih molekula. Ova dinamika može se zamisliti kako slijedi. Počnimo s određenim srednjim zakretnim momentom koji se odražava na dijagramu i karakterizira prisutnost proteina lanca koji je već počeo graditi, pričvršćen na njezinu TRNA i upravo je ušao u ribosoma i povezan s tripletom nove molekule TRNA s odgovarajuća aminokiselina. Očigledno, čin vezivanja TRNA molekule na ribosomu koji se nalazi ribosomski triplet dovodi do takve međusobne orijentacije i bliski kontakt između aminokiselinskog ostatka i proteinskog kruga u konstrukciji, koji se javlja kovalentna veza između njih. Komunikacija se događa na takav način da je kraj proteinskog lanca u izgradnji, na dijagramu pričvršćen na TRNA, prenese se iz ovog TRNA na aminokiselinski ostatak primljenog aminocil-TRNA. Kao rezultat "desnog" TRNA, igrajući ulogu "donatora", bit će slobodna, a protein lanac - "lijevo" (primljeno) aminoacil-TRNA - "lijevo" (primljen) na "Prihvaćeni" - Kao rezultat toga, protein lanac će biti izdužen po aminokiselini i pričvršćen na "lijevo» Trnu. Slijedeći to, "lijevo" TRNA se prenosi zajedno s tripletom koji se odnosi na nukleotid, tada bivši "donor" trna molekula će biti zamijenjena odavde i ostavlja ribosoma, novi TRNA s proteinima koji je bio u izgradnji aminokiselinski ostatak. , a lanac mRNA će se promicati u odnosu na ribosomu na jednom tripletu udesno. Kao rezultat promocije lanca mRNA, jedan triplet u desnoj strani u ribosomu će se pojaviti sljedeći prazan trotić (UOU), a odgovarajući TRNA s aminokiselinom (fenilalanil-TRNA) odmah će biti dodan u skladu s komplementarno načelo. To će ponovno uzrokovati stvaranje kovalentne (peptidne) veze između lanca proteina u konstrukciji i ostatku fenilalanina i slijedi promociju lanca mRNA do jednog tripleta s desne strane sa svim posljedicama koje proizlaze odavde, itd. Na taj se način provode dosljedno, trojku za trojku, povlačeći lanac informacija RNA kroz ribosomu, kao posljedica toga što se lanca IRNN čita "Ribosome u potpunosti, od početka do kraja. U isto vrijeme, postoji dosljedna aminokiselina za aminokiselinu, lanca proteina konjugu. Prema tome, u ribosomu, jedan za drugom dolazi molekule Trvine s aminokiselinama, a molekule TRNA izazivaju bez aminokiselina. Popravak u otopini izvan ribosoma, slobodne molekule TRNA ponovno su povezane s aminokiselinama i ponovno ih nose u ribosomu, čime se ciklički odnosi bez razaranja i promjena.
Glavna figura matrične biosinteze su rNA i DNA nukleinske kiseline , Oni su polimerne molekule koje uključuju dušične baze od pet vrsta, pentoze od dva tipa i ostaci fosforne kiseline. Baze dušika u nukleinskim kiselinama mogu biti purin (adenin, gvanin) i pirimidin (citozin, uracil (samo u RNA), timin (samo u DNA)). Ovisno o strukturi ugljikohidrata, označite ribonukleinske kiseline - sadrži Ribosa (RNA) i deoksiribonukleinske kiseline - sadržavati dezoksiribozu (DNA).
Pojam " matrična biosinteza"To podrazumijeva sposobnost ćelije da sintetizira polimerne molekule, kao što je nukleinske kiseline i proteini, na temelju predloška - matrica. Time se osigurava točan prijenos najsloženije strukture od već postojećih molekula do novoizmjenjivih.
Glavni postulat molekularne biologije
U velikoj većini slučajeva prijenos nasljednih informacija iz majčinske stanice na podružnicu provodi se pomoću DNA (replikacija). Da biste koristili genetske informacije, samostalna stanica zahtijeva RNA formirana na DNA matrici (transkripcija). Nadalje, RNA je izravno uključena u sve faze sinteze proteinskih molekula (prijevod), osiguravajući strukturu i aktivnost stanice.
Na gore navedeno središnja molekularna biologijaPrema kojem se prijenos genetskih informacija provodi samo na nukleinskoj kiselini (DNA i RNA). Primatelj informacija može biti druga nukleinska kiselina (DNA ili RNA) i proteina.