Tema prelegerii noastre "Funcțiile coaja de emisfere mari. "
Creierul este în cavitatea craniului creierului. Are suprafețe convexe topless și inferioare, precum și o suprafață compusă - baza creierului.
Biroul mare constă din două emisfere - dreapta și stânga, care sunt legate de un corp de porumb de comisiu. Dreptul de emisfere dreapta și stâng sunt împărțite cu decalajul longitudinal. În cadrul Comisiei, există o seif, care este două colanți fibroase curbe, care în partea de mijloc sunt interconectate și în față și spate, formând stâlpi și picioare ale arcului. Frontul din coloanele arcului este spike-ul din față. O placă subțire de țesut creierului subțire este întinsă între corpul de porumb și partiția arc-transparentă.
Emisferele au suprafețe superioare, medii și inferioare. Superior-latheral - convex, medial - plat, cu care se confruntă aceeași suprafață a unei alte emisfere și o formă neregulată mai mică. Pe trei suprafețe sunt caneluri adânci și mici, iar între ele sunt girus. Brazde - aprofundarea între convoluții. Înfășurarea - creșterile creierului.
Suprafețele emisferelor creierului mare sunt separate unul de celălalt de margini - partea de sus, novenerală și nouăntică. În spațiul dintre cele două emisfere, un proces mare în formă de bolnavă include o placă subțire de o coajă solidă, care pătrunde într-o fantă longitudinală a unui creier mare, fără a ajunge la un corp de porumb și separă emisfera dreaptă și stângă una de cealaltă .
Cele mai proeminente secțiuni ale emisferelor au fost numite stalpi: frontal, occipient și temporal. Suprafețele de relief ale emisferelor creierului mare sunt foarte complexe și conexiuni cu prezența unei brazde mai mari sau mai puțin adânci a unui creier mare și situată între ele cu înălțimi în formă de rume. Adâncimea, lungimea unor brazde și convoluțiile, forma și direcția lor sunt foarte variabile.
Fiecare emisferă este împărțită în acțiuni - frontală, întunecată, occipientă, temporală, insulă. Brăzda centrală separă cota frontală de la parietal, canelura laterală - cota temporală de la frontal și întuneric, canelura întunecată-occipientă împărtășește nuanțele întunecate și occipale.
Emisferele mari constă în straturi orizontale situate în direcția de pe suprafața adâncă.
I. Stratul molecular are celule foarte mici, dar un număr mare de dendrite de ramificare a celulelor piramidale care formează plexul situat paralel cu suprafața. La aceste dendrite, sinapsele fibrelor aferente provenind din nucleele Talamus asociative și nespecifice sunt formate.
II. Stratul de grainiu exterior este compilat în principal de stereo și celule piramide parțial mici. Fibrele celulare ale acestui strat sunt situate în principal de-a lungul suprafeței cortexului, formând conexiuni corticocortice.
III. Stratul de piramidă exterioară constă în principal din celulele piramidale de dimensiuni medii. Axele acestor celule, precum și stratul de celule granulare II, formează conexiuni asociative corticocortice.
IV. Stratul de granulare interior în funcție de natura celulelor și localizarea fibrelor lor este similar cu stratul exterior de cereale. Pe neuronii acestui strat, terminațiile sinaptice ale fibrelor aferente provenind din neuronii nucleelor \u200b\u200bspecifice ale talamusului și, prin urmare, din receptorii sistemelor senzoriale.
V. Stratul de piramidă interioară este format din celulele piramide medii și mari, iar celulele piramide gigantice ale BETZ sunt situate în crusta în mișcare. Axonii acestor celule formează autostrăzi corticospinale și corticobulble efectiv.
VI. Stratul celulelor polimorfe este predominant celule asemănătoare coloanei vertebrale ale căror axe formează căi corticotice.
În primul și al patrulea strat există percepția și prelucrarea semnalelor care intră în coajă. Neuronii din al doilea și al treilea straturile efectuează conexiuni asociative corticocortice. Căile eferente care părăsesc gaura se formează în principal în straturile a cincea și a șasea.
Diviziunea mai detaliată a scoarței în diverse domenii a fost efectuată pe baza formei și localizării neuronilor Brodman, care a alocat 11 zone, inclusiv 52 de câmpuri, dintre care multe se caracterizează prin caracteristici funcționale și neurochimice. Potrivit lui Brodman, zona frontală include următoarele câmpuri: 8, 9, 10, 11, 12, 44, 45, 46, 47. În regiunea prezentatorului, aceasta include câmpul 4 și 6, regiunea post-centrală include 1, 2, 3 și 43. Zona parietală include câmpurile 5, 7, 39, 40 și regiunea occipitală 17 18 19. Zona temporală constă dintr-un număr foarte mare de domenii.
În zonele de cortex, senzoriale, asociative și motoare se disting, pe baza locației neuronilor:
Problema localizării funcțiilor în crusta de emisfere mari are trei concepte:
Principiul localizării înguste - toate funcțiile sunt plasate într-o structură luată separat.
Conceptul de echipamente - diferite structuri corticale sunt egale funcționale.
Principiul multifuncționalității domeniilor corticale.
Proprietatea multifuncționalității permite ca această structură să fie inclusă în furnizarea de diferite forme de activitate, în timp ce punerea în aplicare a funcției de bază, inerente genetic. Gradul de multifuncționalitate a diferitelor structuri corticale ale non-Einakov: de exemplu, în câmpurile de coajă asociativă, este mai mare decât în \u200b\u200bcâmpurile senzoriale primare și în structurile corticale mai mari decât în \u200b\u200btulpină. În centrul multifuncționalității se află admiterea multi-canal la coaja creierului excitației aferente, suprapunerea excitațiilor aferente, în special asupra nivelurilor thalamice și corticale, influența modulativă a diferitelor structuri (talamus nespecifică, ganglionii bazali) asupra funcțiilor corticale , interacțiunea tractului cortical-subcortic și de interpionie al excitației.
Una dintre cele mai mari opțiuni pentru separarea funcțională a noului cortex al creierului este selecția regiunilor senzoriale, asociative și motorii în ea.
Zonele de atingere ale cortexului sunt zone în care sunt proiectate stimuli senzoriali. Zonele senzoriale ale cortexului sunt numite altfel: Departamentele de proiecție sau cu șuruburi de analizoare. Acestea sunt situate în principal în acțiuni întunecate, temporale și occipale. Căile aferente din bor senzoriale provin în principal din nucleele senzoriale specifice ale talamusului. Coaja senzorială are straturi bine pronunțate și a patra și se numește granulară.
Zonele cortexului senzorial, iritarea sau distrugerea cărora cauzează schimbări clare și constante în sensibilitatea organismului, se numesc zone primare senzoriale. Acestea constau în principal din neuroni monomode și de senzații de aceeași calitate. În zonele senzoriale primare, există, de obicei, o reprezentare spațială clară a părților corpului, a câmpurilor de receptor. Există zone senzoriale secundare mai puțin localizate în jurul zonelor senzoriale primare, ale căror neuroni polimodali răspund la mai mulți stimuli.
Cea mai importantă zonă senzorială este coaja întunecată a înfășurării post-centrale și partea corespunzătoare a felii de paravole pe suprafața medială a emisferelor, care este indicată ca regiunea primară somatosensorică. Există o proiecție a sensibilității pielii din partea opusă a corpului de tactil, durere, receptori de temperatură, sensibilitate interoceceptivă și sensibilitate a sistemului musculoscheletal de la receptorii musculari, articulari și de tendon. Proiecția secțiunilor corpului în această zonă se caracterizează prin faptul că proiecția capului și a părților superioare ale corpului este situată în scăderile inferioare ale înfășurării post-centrale, proiecția jumătății inferioare a corpului și Picioarele - în zonele nucleare superioare ale înfășurării, proiecția piciorului inferior și oprirea - în miezul felii de parackentrale de pe suprafața medială a emisferelor. În același timp, proiecția celor mai sensibile situri (limbaj, buze, laringe, degete) are zone relativ mari în comparație cu alte părți ale corpului. Se presupune că, în zona de sensibilitate tactilă a limbii, există și o proiecție a sensibilității gustului.
Regiunea somatosensorică secundară de dimensiuni mai mici este situată pe peretele superior al canelurii laterale, la marginea intersecției sale cu brazdele centrale. Funcțiile regiunii somatosensoriale secundare sunt studiate prost. Se știe că localizarea suprafeței corpului este mai puțin clară, impulsarea vine aici atât din partea opusă a corpului, cât și din partea "lui", sugerează participarea sa la coordonarea senzorială și a motorului laturile corpului.
O altă zonă senzorială primară este coaja de auz, care este situată în adâncurile brazdei laterale. În această zonă, ca răspuns la iritarea receptorilor de audit ai organului Cortiev, se formează senzațiile de sunet, schimbând volumul, tonul și alte calități. Are o proiecție topică clară: în diferite părți ale cortexului, sunt prezentate diferite secțiuni ale organului Cortiyev. Miezul de proiecție al acțiunii temporale include, de asemenea, centrul analizorului vestibular în convingerile temporale superioare și medii. Informațiile senzoriale prelucrate sunt utilizate pentru a forma o "schemă corporală" și reglementează funcțiile cerebelului (traseul temporal-bridge).
O altă zonă de proiecție primară a noii coajă este situată în cortexul occipital - regiunea vizuală primară. Aici are o reprezentare locală a receptorilor retinei și fiecare punct al retinei corespunde secțiunii sale din coaja vizuală, în timp ce zona locului galben are o zonă mare a reprezentării. Datorită intersecției incomplete ale tractului vizual în regiunea vizuală a fiecărei emisfere, aceleași vârste ale retinei sunt proiectate. Prezența în fiecare emisferă a proiecției retinei a ambelor ochi este baza viziunii binoculare.
O iritație a crustei câmpului al 17-lea duce la apariția senzațiilor de lumină. Lângă câmpul 17 este coaja regiunii vizuale secundare. Neuronii acestor zone sunt polimodali și răspund nu numai la lumină, ci și pe stimulii auditivi, auditivi. În această regiune vizuală, se produce sinteza diferitelor tipuri de sensibilitate și imaginile vizuale mai complexe și apare identificarea acestora. Iritarea acestor câmpuri determină halucinații vizuale, senzații obsesive, mișcări ale ochilor.
Partea principală a informațiilor despre mediu și a mediului interior al corpului, care a intrat în borul senzorial este transmisă pentru prelucrarea ulterioară în coaja asociativă.
Zonele cortexului asociativ includ secțiuni ale unei coaja noi situate în apropierea zonelor de atingere și motoare, dar care nu funcționează direct de funcții sensibile și motorii. Limitele acestor zone nu sunt indicate suficient, incertitudinea se referă în principal la zonele de proiecție secundară, ale căror proprietăți funcționale sunt tranzitorii între proprietățile proiecției primare și zonele asociative. La om, coaja asociativă este de 70% din neocortex.
Principala caracteristică fiziologică a neuronilor cortexului asociativ este polimodalitatea: ei răspund la mai mulți stimuli cu aproape aceeași forță. Polimodalitatea (polyessence) a neuronilor din coaja asociativă este creată din primul rând, prezența legăturilor corticocortice cu diferite zone de proiecție, în al doilea rând, datorită principalei intrare aferentă din nucleele de talamus asociative, în care a existat deja dificil Prelucrarea informațiilor din diferite moduri sensibile. Drept urmare, coaja asociativă este un aparat puternic al convergenței diferitelor excitații senzoriale, ceea ce face posibilă o prelucrare complexă a informațiilor despre mediul extern și intern al corpului și îl folosește pentru implementarea unui psiho-fiziologic mai mare Funcții. În cortexul asociativ, se disting trei sisteme creierului asociativ: Talamotimen, Talamolobnye și TalamumoChnaya.
Sistemul Talamum este reprezentat de zonele asociative mai întunecate, care primesc principalele intrări aferente din partea din spate a nucleelor \u200b\u200bde talamus asociative. Scoarța asociativă întunecată are randamente eferente asupra kernel-urilor de talamus și hipotalamus, orificiul motorului și miezul sistemului extrapiramidal.
Principalele funcții ale sistemului de talamum sunt gnoza, formarea "schemei corpului" și a praxei.
Sub gnoza înțelege funcția diferitelor tipuri de recunoaștere: formulare, valori, valori ale obiectelor, înțelegerea discursului, cunoașterea proceselor, modelele. Funcțiile gnostice includ o evaluare a relațiilor spațiale. În cortexul parietal, centrul stereogenis se distinge din spatele departamentelor medii ale bobinei post-centrale (câmpurile 7, 40, parțial 39) și asigurând capacitatea de a recunoaște obiectele la atingere. O variantă a funcției gnostice este să se formeze în conștiința unui model de corp tridimensional, a cărei centru este situat în câmpul 7 din coaja parietală. Lângă Praxis. Înțelegeți acțiunea vizată, centrul se află în urinețul Suprava (câmpurile 39 și 40 din emisfera dominantă). Acest centru oferă stocarea și implementarea programului de acte automate de motor.
Sistemul tamulobular este reprezentat de zone asociative ale unei scoarțe frontale având o intrare aferentă fundamentală din miezul asociativ al aparatului mediu al talamusului. Funcția principală a cortexului asociativ frontal este formarea programelor de comportament concentrat, în special într-o nouă situație pentru o persoană. Implementarea acestei funcții generale se bazează pe alte funcții ale sistemului tamulobic: 1) formarea unei motivații dominante care asigură direcția comportamentului uman. Această funcție se bazează pe legături bilaterale apropiate ale Liniei cu un sistem limbic și rolul acestuia din urmă în reglementarea emotiilor mai mari ale unei persoane legate de activitățile sale sociale și creativitatea; 2) furnizarea de predicție probabilistă, care este exprimată prin schimbarea comportamentului ca răspuns la schimbările în mediul de mediu și la motivația dominantă; 3) auto-controlul acțiunii prin compararea constantă a rezultatului acțiunii cu intențiile inițiale, care este asociată cu crearea asistenței de predicție (acceptorul de rezultate).
În cazul deteriorării cortexului fronforțional frontal, unde sunt intersectați conexiunile dintre cota frontală și talamus, persoana devine brută, insensibilă, nesigură, el are tendința de a repeta orice acte motorii, deși situația sa schimbat deja și Trebuie efectuate alte acțiuni.
Sistemul de examinare thalamic nu este suficient de studiat. Dar dacă vorbim despre cortexul temporal, atunci trebuie remarcat faptul că unele centre asociative, cum ar fi stereoognoza și praxis, includ zonele cortexului temporal. În cortexul temporal există un centru de vorbire de vorbire, situat în secțiunile din spate ale înfășurării temporale superioare. Acest centru oferă gnoza de vorbire - recunoașterea și depozitarea discursului oral, ambele proprii și ale altora. În partea de mijloc a înfășurării temporale superioare, există un centru de recunoaștere a sunetelor muzicale și a combinațiilor lor. La granița fracțiunii temporale, întunecate și occipitale este centrul de lectură, oferind recunoașterea și stocarea imaginilor scrise.
În cortexul motorului, regiunile principale și secundare sunt distinse.
În cortexul principal al motorului, neuronii sunt situați, inervând mișcările mușchilor feței, corpului și membrelor. Are o proiecție topografică clară a mușchilor corpului. În același timp, proiecțiile mușchilor extremităților inferioare și a trunchiului sunt situate în secțiunile superioare ale unei consolidări prezennte și ocupă o zonă relativ mică, iar proiecția mușchilor extremităților superioare, a fețelor și a limbii sunt situate în secțiunile inferioare ale înfășurării și ocupă mai mare. Modelul principal al reprezentării topografice este că reglementarea activității musculare care furnizează mișcările cele mai exacte și variate (vorbire, scrisoare, expresii faciale) necesită participarea marilor în zona de cortex de mișcare. Reacțiile motorului la iritarea cortexului principal de motor sunt efectuate cu un prag minim (excitabilitate ridicată) și sunt reprezentate de abrevieri elementare ale mușchilor din partea opusă a corpului (pentru că mușchii abreviere a capului poate fi bilaterală). Sub înfrângerea acestei zone a cortexului, capacitatea de a pierde mișcările coordonate subtile a mâinilor este pierdută, în special degetele.
Cortexul secundar de mișcare este situat pe suprafața laterală a emisferelor, înainte de înfășurarea precentrală. Ea efectuează funcții mai mari ale motorului legate de planificarea și coordonarea mișcărilor arbitrare. Coaja câmpului 6 primește partea principală a impulsurilor eferente de nuclee bazale și cerebelul și participă la transcodarea informațiilor despre programul de mișcări complexe. Iritarea de scoarță a câmpului 6 provoacă, de exemplu, mișcări mai complexe coordonate, transformând capul, ochiul și trunchiul în partea opusă, contracțiile prietenoase ale mușchilor-flexor sau extensori de mușchi pe partea opusă. În Primorny Core, există centre de motoare asociate cu funcțiile sociale ale unei persoane: centrul discursului scris în partea din spate a înfășurării frontalelor medii, centrul scurgerii motorului Brock în partea din spate a sediului inferior, Furnizarea de praxis de vorbire, precum și un centru de motor muzical care determină tonalitatea discursului, abilitatea de a cânta.
Cortexul motorului este mai bun decât în \u200b\u200balte zone ale scoarței, se exprimă un strat care conține celulele piramide uriașe ale BETZ. Neuronii de cortex ai motorului primesc intrări aferente prin talamus din receptorii musculari, articulari și de piele, precum și de la nucleele bazale și cerebelul. Principala ieșire efectivă a cortexului motor pe centrele de motor stem și spinale formează celule piramidale ale celui de-al cincilea strat. Piramida și neuronii de inserție conjugată sunt localizați pe verticală față de suprafața cortexului și formează difuzoarele motorului neuronal. Neuronii piramidali ai coloanei motorului pot excita sau frâna tulpinile și centrele spinării. Coloanele învecinate din planul funcțional se suprapun, iar neuronii piramidali care reglementează activitatea unui mușchi sunt de obicei localizați nu într-una, ci în mai multe coloane.
Principalele conexiuni efective ale cortexului motor sunt efectuate prin căile piramidale și extrapiramidale, care încep cu celulele piramide gigantice ale pariului și celulele piramide mai mici ale celui de-al cincilea strat al crustei cortexului de prezentare (60% din fibre) , coaja de premotor (20% fibre) și înfășurarea post-centrală (20% fibre). Celulele mari piramidale au o axonă rapidă și o activitate de impuls de fundal de aproximativ 5 Hz, ceea ce crește la 20-30 Hz la conducere. Aceste celule inervați -Motoniile mari (de mare viteză) în centrele motor ale trunchiului și măduvei spinării care reglementează mișcările fizice. Axoanele de mielină subțire de mielină pleacă de la celulele mici piramidale. Aceste celule au o activitate de fundal de aproximativ 15 Hz, ceea ce crește sau scade în timpul conducerii. Ei inervați mici (low-laminate)? -Motoneons în centrele de motor stem și spinale, reglarea tonului muscular.
Căile piramide constau din 1 milion de fibre ale căii corticospinale, care încep cu cortexul treimii superioare și mijlocii din scufundarea prezentatorului și 20 de milioane de fibre ale căii Corticobulbar, care începe cu cortexul treimii inferioare a prezenței ispud.
Fibrele calea piramidală se termină pe alfa-motoanele nucleelor \u200b\u200bmotorului din al treilea - al șaptelea și al nouă doisprezece nervi cranieni (calea corticobulbulberry) sau pe centrele de motor spinale (calea corticospinală).
Prin intermediul motorii și al piramidelor, sunt efectuate mișcări simple arbitrare și programe complexe de motoare vizate, de exemplu, abilitățile profesionale, formarea cărora începe în ganglionii bazali și cerebelul și se termină în crusta motorului secundar.
Majoritatea fibrelor căilor piramidale efectuează Crucea, cu toate acestea, o mică parte a fibrelor rămâne necombală, ceea ce contribuie la compensarea funcțiilor de mișcare cu leziuni cu leziuni unilaterale. Prin căile piramidale, acesta oferă funcțiile și coaja premotorului: abilitățile motorii, cap de cap, ochi și tors în direcția opusă, precum și discurs. În reglementarea scrisorii și în special a discursului verbal, există o asimetrie pronunțată a emisferelor mari ale creierului: în 95% din partea dreaptă și 70% din discursul stâng este controlat de emisfera stângă.
Căile extrapiramidale corticale includ căile corticrurale și corticorculare începând cu aproximativ acele zone care dau începutul căilor piramidale. Fibrele căii corticorulare se termină pe neuronii nucleei roșii din creierul mijlociu, de la care se duc regulile ruble.
Fibrele căilor corticorculare se termină pe neuronii miezurilor mediale ale formării reticulare a podului (sunt urmate căile reticulospinale mediane) și pe neuronii miezurilor gigalide reticulare ale creierului alungit, din care începe rutele reticulospinale laterale.
Prin aceste căi sunt reglementate de tonuri și posesiuni care oferă mișcări precise vizate. Calea extrapiramidală corespondentă sunt componenta sistemului creierului esseramidic la care cerebelul, ganglionii bazali, centrele motoare ale cilindrului. Sistemul extrapiramine îndeplinește tonul, postura de echilibru, execuția actelor motor explodate, cum ar fi mersul pe jos, alergarea, vorbirea, scrisoarea. Deoarece căile de corticopiramid dau numeroasele lor structuri colaterale cu un sistem extrapiramidal, ambele sisteme funcționează în unitate funcțională.
Evaluarea rolului diferitelor structuri ale capului și măduvei spinării în termeni generali în reglarea mișcărilor complexe direcționale, se poate observa că mișcarea (motivația) mișcării este creată în sistemul limbic, planul de mișcare este în Cortexul asociativ de emisfere mari, programe de mișcări - în ganglionii bazali, cerebelul și cortexul premotor și performanța mișcărilor complexe apare printr-un motor, centre de motoare ale cilindrului și măduvei spinării.
Relația intermetroasă la oameni se manifestă în două forme - asimetria funcțională a emisferelor mari și a activităților lor comune.
Asimetria intereselor ca una dintre caracteristicile importante ale funcționării celor mai înalte departamente ale creierului este determinată în principal de două momente: 1) localizarea asimetrică a aparatului nervos al celui de-al doilea sistem de semnalizare și 2) dominația mâinii drepte ca un mijloc puternic de Adaptiv comportament adaptiv uman. Acest lucru explică faptul că primele idei despre rolul funcțional al asimetriei intermetroase au apărut numai atunci când a fost posibil să se stabilească localizarea centrelor de vorbire nervoasă (motor-centru de brock și senzorial - centrul Wernik în emisfera stângă).
Proiecția încrucișată a speciilor de sensibilitate senzorială și piramide în aval - regulatoare ale sferei motorului corpului - în combinație cu localizarea spre stânga a centrului de discurs orală și de scriere, determină rolul dominant al emisferei stângi în comportamentul unei persoane gestionate de crusta de emisfere mari.
Datele experimentale obținute confirmă ideea rolului dominant al emisferei stângi a creierului în implementarea funcțiilor celui de-al doilea sistem de semnalizare, în operațiunile de gândire, în activități creative cu predominanța formelor de gândire abstractă. În general, se poate presupune că persoanele cu dominanță de uscare a frunzelor aparțin tipului de gândire și cu dominația dreaptă - la artistice.
Asimetria funcțională a emisferelor este cea mai importantă proprietate psihofiziologică a creierului uman. Mental, senzorial și motor interplassar asimetria funcțională a creierului este izolat.
În studiul funcțiilor psihofiziologice, sa arătat că în vorbire, canalul de informații verbale este controlat de emisfera stângă, iar canalul non-pepolit (voce, intonație) are dreptate.
Gândirea abstractă și conștiința sunt conectate, în principal cu emisfera stângă. La dezvoltarea unui reflex condiționat în faza inițială, emisfera dreaptă domină și în timpul consolidării reflexului - stânga.
Hemisfera dreaptă efectuează prelucrarea informațiilor în același timp, sintetic, în conformitate cu principiul deducerii, semnele spațiale și relative ale subiectului sunt mai bine percepute. Emisfera stângă produce procesarea informațiilor secvențial, analitic, pe principiul inducției, semnele absolute ale subiectului și relația temporară percepe.
În sfera emoțională, emisfera dreaptă determină emoțiile în principal negative, controlează manifestările emoțiilor puternice, în general, este mai "emoțional". Emisfera stângă determină cele mai multe emoții pozitive, controlează manifestarea emoțiilor mai slabe.
În sfera senzorială, rolul emisferelor drepte și stângi se manifestă cel mai bine cu percepția vizuală. Emisfera potrivită percepe imaginea vizuală Hivo, imediat în toate detaliile, este mai ușor rezolvă problema distingerii obiectelor și identificarea imaginilor vizuale ale obiectelor, ceea ce este dificil de a descrie în cuvinte, creează premisele gândirii senzuale concrete. Emisfera stângă estimează imaginea vizuală dezmembrată, analitic, fiecare semn analizat separat. Subiecții familiari sunt mai ușor și sarcinile similarității obiectelor sunt rezolvate, imaginile vizuale sunt lipsite de detalii specifice și au un grad ridicat de abstractizare; Creați medii de gândire logică.
Asimetria motorului este exprimată în primul rând, în primul rând, în dreapta-leopard-leopard, care este controlată de motocicleta de emisferă opusă. Asimetria altor grupuri musculare are o persoană și nu specii.
Paritatea în activitățile de emisfere mari este asigurată de prezența unui sistem comandat (organism de porumb, al Comisiei anterioare și spate, hipocampale și hipenulare, lupta interstigalizată), care combină anatomic două emisfere cerebrale. Cu alte cuvinte, ambele emisfere sunt conectate nu numai prin conexiuni orizontale, ci și verticale.
Principalele fapte obținute prin tehnicile electrofiziologice au arătat că excitația unei singure emisfere din secția de iritare este transmisă prin sistemul de punere în funcțiune nu numai în partea simetrică a unei alte emisfere, ci și în secțiunile asimetrice ale scoarței. Studiul metodei reflexelor condiționate a arătat, în curs de dezvoltare a unui reflex, "transferul" unei conexiuni temporare la o altă emisferă. Formele elementare ale interacțiunii a două emisfere pot fi efectuate prin cvadruses și formarea reticulară a cilindrului.
O persoană, ca multe animale, majoritatea organelor sunt asociate: două mâini, două picioare, două ochi, două urechi, două rinichi, două emisfere ale creierului. Paritatea organelor nu înseamnă funcționarea lor identică. Știm ce mână avem conducerea - efectuează cele mai complexe și subtile operațiuni. Majoritatea oamenilor au o mână dreaptă. Mâncăm, coasem, scriem, trageți mâna dreaptă. Printre oameni - cu mâna dreaptă care utilizează dreptaci pentru acțiune exactă, 90%, în timp ce au rămas în medie 10%.
Stângi de toate rasele și culturile din trecut și prezentul au fost într-o minoritate printre mediul potrivit.
La studierea problemei originii leficăriei, s-au distins trei direcții principale: "genetice", "culturale" și "patologice".
În prezent, două modele genetice au primit cea mai mare distribuție. Conform unei asimetrice cerebrale, este determinată prezența unei gene, numită-o de factorul "schimbării drepte". Dacă acest factor este disponibil într-un individ, acesta din urmă este predispus să fie dreptaci. Dacă nu există nici un factor, o persoană poate fi fie stângaci, fie chiar în funcție de circumstanțe aleatorie. În acest caz, leziunile cerebrale în perioada postnatală prenatală și timpurie, care poate afecta realizarea fenotipică a factorului "Shift dreapta".
Un model mai complex a fost propus de Levi și Nagilaki (1972). Acești oameni de știință sugerează că arbitrul este o funcție a două gene. O genă cu două alele determină emisfera care va controla vorbirea și mâna de conducere.
Probabilitatea de non-genetica, dar citoplasmatica codificată de asimetrie indică Morgan (1978), punând în continuare un concept conform căruia lateralizarea creierului și preferințele manuale sunt considerate într-un aspect comercial larg. Se presupune că dezvoltarea creierului este sub influența gradientului stâng-drept, ceea ce duce la o maturare anterioară și rapidă în ontogeneza emisferei stângi, care, în același timp, are un efect inhibitor asupra dreptului - Ca urmare, există dominanță a emisferei stângi asupra discursului și drepturilor.
Cu direcția "genetică" combină în mod direct cercetarea legată de identificarea stigmaticii anatomice, fiziologice și morfologice, caracteristice cu mâna dreaptă și cu mâna stângă. Se arată că în partea dreaptă Silvieva, canelura din dreapta este situată deasupra stângii, în timp ce 71% din brațele stângi și stângi sunt aproximativ simetrice.
Bunăstarea corectă Există un diametru mai mare al arterei carotide interioare din stânga și deasupra presiunii în ea decât în \u200b\u200bdreapta, iar partea stângă este o imagine opusă.
Disocierea similară este detectată în partea dreaptă și stângaci atunci când studiază artera cerebrală medie. Ipoteza Gershvinda și Galaburd implică, de asemenea, un efect endocrin asupra formării diferențelor în structura creierului bărbaților și femeilor. Teoria previne este cunoscută, conform căreia lateralizarea cerebrală la om se formează cu dezvoltarea prenatală asimetrică a sistemului interior al urechii și al labirintului.
Există, de asemenea, o ipoteză genetică și culturală a asimetriei funcționale. Omul de știință englez din Lalland Cambridge și colegii săi consideră că levcy este determinată la fel de genetic și cultural.
Alternativă "genetică" sunt ipotezele apariției asimetriei intermetrack, bazate pe recunoașterea determinării rolului condițiilor culturale în formarea opresiunii. Conceptele "culturale și sociale" au în vedere Levierty Cutenție ca o consecință a educației sociale, a experienței, a condițiilor de viață.
Împreună cu teoriile prezentate mai sus, ideile despre originea patologică a Lefta sunt larg răspândite. Un punct extrem de vizualizat aderă la Bankan (1973), care susține că orice manifestare a stângii este o consecință a unui prejudiciu generic. Potrivit lui Chuprikova (1975), schimbarea dominanței motorii este una dintre dovezile obiective ale encefalopatiei congenitale. În confirmare, se administrează faptele de creștere a celor stângaci între gemeni, ale căror caracteristici de dezvoltare prenatală implică riscul hipoxiei creierului intrauterin. În favoarea acestei abordări, rezultatele testelor WADA, conform cărora daunele la emisfera stângă în stadiile incipiente ale ontogenezei pot duce la o schimbare a brațului de conducere și a emisferei dominante în vorbire.
Studiul originii lateralității continuă. Abundența faptelor, uneori contrazice reciproc, arată că fiecare dintre teoriile asimetriei funcționale inter-imparale a creierului necesită o fundamentare suplimentară. În același timp, este evident că principiile fundamentale ale abordărilor de mai sus reprezintă baza pentru viitorul studiu sistemic, necesitatea care rezultă din multe probleme și probleme rămase deschise.
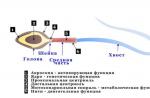
Funcțiile cortexului de emisfere mari
În funcționalitatea emisferelor mari de a împărtăși în trei domenii: senzoriale, motor (motor) și coajă asociativă. Zona senzorială include acele zone ale cortexului de emisfere mari, în care se preconizează stimulii senzoriali. Coaja senzorială este situată în principal în fragmentele întunecate, temporale și occipale ale creierului mare. Căile aferente din bor senzoriale provin în principal din nucleele senzoriale specifice ale talamusului. Touch Cortex Zone includ zonele primare și secundare latra. În zonele primare, sentimentele de aceeași calitate. În zone secundarecorams formează senzații care apar ca răspuns la acțiunea mai multor stimuli.
Principala zonă senzorială a coajului este în:
Urgenul post-central: sensibilitatea pielii de la receptorii tactili, de temperatură a durerii; Sensibilitatea sistemului musculoscheletal - mușchi, articulații, tendoane; Tactilă și sensibilitatea gustului limbii.
- convoluție temporală medie (și. Geshlya), senzațiile de sunet sunt formate aici -
Convulsia temporală superioară și medie, centrul analizorului vestibular este localizat aici, se formează senzațiile de "schema corpului".
- regiuneaClinovoidul este o zonă vizuală primară situată în nucleul occipital.
Zona asociativă a cortexului include zone situate în apropierea zonelor senzoriale și de motor, dar nu efectuează funcții sensibile sau motorii direct. Limitele acestor regiuni nu sunt indicate bine. În kretul asociativ, puteți evidenția zonele:
Sistemul Tamolas;
Talamum System;
Sistemul talamumian.
Sistemul tamolobic este implicat în formarea motivării dominante: această funcție se datorează unei legături bilaterale între coaja frontală și sistemul limbic, asigură probabilitățile de predicție și auto-controlul acțiunii prin compararea constantă a rezultatului acțiunii cu intențiile inițiale .
Sistemul de talamum îndeplinește funcțiile gnozei, formarea "schemei corpului" - stereoomismul și praxisul. Gnoza este o funcție a diferitelor tipuri de recunoaștere: formulare, valori, valori ale obiectelor, înțelegerea discursului, cunoașterea proceselor și modelelor. Funcția stereonisis asigură capacitatea de a recunoaște elementele la atingere. În centrul stereoognisi, senzațiile sunt formate responsabile pentru crearea unui model de corp tridimensional - "Schema corpului". Praxis este o funcție care vizează efectuarea oricărei activități, centrul său este situat în Urinetul Naparase, asigură depozitarea și implementarea programului de acte motorii (strângere de mână, pieptății etc.).
Sistemul talamumian În înfășurarea superioară a cortexului temporal, există un centru de vorbire de vorbire de către Wernik. Oferă gnoza de vorbire - recunoașterea și stocarea discursului oral. În partea de mijloc a bobinei temporale superioare este centrul recunoașterii sunetelor muzicale. În limitele fracțiunii temporale, întunecate și occipitale, există un centru scris de citire a vorbirii, oferind recunoașterea și stocarea imaginilor scrise.
Motorul motorului ocupă zona lobului frontal al crustei de emisfere mari. LA coreea de motor primară (Crucea prezentală) Există neuroni, înnegrirea mișcărilor mușchilor feței, a torsului și a membrelor. Cord secundar de mișcare Situat pe suprafața laterală a emisferelor, înainte de scufundarea prezentatorului (coaja premotorului). Ea efectuează funcții mai mari ale motorului legate de planificarea și coordonarea mișcărilor arbitrare. Această coajă primește partea principală a impulsului eferent de la nucleele bazale și cerebelul și participă la transcodarea informațiilor despre programele de mișcări complexe. Există centre asociate cu funcțiile sociale ale unei persoane în Core Premotor:
În partea din spate a bobinei frontale medii - centrul discursului scris,
În partea din spate a ferestrelor frontale inferioare, centrul discursului motorului Brock-ului, oferind libertatea de vorbire, precum și un centru de motor muzical care determină tonalitatea discursului.
Neuronii de cortexi ai motorului primesc intrări aferente prin talamus de la receptori musculari, articulari și de piele, precum și de la ganglionii bazali și cerebelul. Principalele rezultate ale cortexului motor pe centrele de motor stem și spinale formează celule piramidale ale crustei. Neuronii piramidali ai cortexului motorului excită sau frânează tulpina și centrele spinării.
Unul dintre principiile de bază ale funcționării crustei de emisfere mari ale creierului este principiul asimetriei inter-impar. Asimetria intermetroasă se datorează localizării asimetrice a aparatului nervos al celui de-al doilea sistem de semnal și dominanța mâinii drepte, ca mijloc de comportament adaptiv. Conform neurofiziologiei moderne (V.L. Bianci), emisfera la jumătatea creierului stâng la oameni se specializează în realizarea funcțiilor simbolice verbale și emisfera corectă asupra implementării funcțiilor figurative spațiale. Rezultatul unei astfel de separare funcționale este asimetria activității mentale, care se manifestă prin diferențe în tipurile de operații mentale. Dominanța emisferei stângi determină tipul de gândire și emisfera dreaptă de gândire artistică.
MUNCA PRACTICA
Pentru a determina coeficientul de asimetrie funcțional, se utilizează formulare, care sunt foi de hârtie (A4), pe care 8 dreptunghiuri egale sunt amplasate la 4 la rând. Fiecare dreptunghi este umplut în mod constant de la stânga la dreapta de la # 1 până la numărul 4 și în direcția opusă de la numărul 5 până la numărul 8. Forma formularului este prezentată în figura 1.
Figura 1 - Jobul gol
Instrucțiuni: "În semnalul meu, trebuie să începeți să ridicați punctele din fiecare dreptunghi al formularului. Pentru timpul stabilit pentru fiecare dreptunghi (5 secunde), trebuie să puneți în ea ca puncte mai mari. Du-te de la un dreptunghi la altul de care aveți nevoie pentru a comanda, fără a întrerupe lucrările. Tot timpul când lucrați la tempo maxim. Acum luați un creion în dreapta (sau mâna stângă) și puneți-l în fața primului dreptunghi al formularului. "
În opritor, experimentatorul dă un semnal: "a început!", Apoi la fiecare 5 secunde dă comanda: "Următorul!". După 5 secunde de lucru într-un dreptunghi nr. 8, experimentatorul prezintă comanda: "Stop". Calculați numărul de puncte din fiecare pătrat și completați tabelul 1 în notebook-ul de lucru.
Tabelul 1 - Protocolul de cercetare
Folosind rezultatele tabelului 1, efectuați un grafic al relației dintre timpul de execuție a timpului (Axa X) și numărul de puncte pentru fiecare mână (axa Y). Luați o concluzie, ghidată de următorul model: mâna dreaptă este performanța mâinii drepte deasupra operabilității lui Lefters, iar partea stângă este opusă.
Calculați coeficientul de asimetrie funcțională asupra operabilității mâinii stângi și drepte, obținând valorile totale ale capacității de lucru prin adăugarea tuturor datelor pentru fiecare dintre cele opt dreptunghiuri. Pentru a calcula, utilizați formula pentru a estima coeficientul de asimetrie funcțională (1):
KF A \u003d \u200b\u200b[(SR - SL) / (SR + SL)] (1)
unde KF A este coeficientul de asimetrie funcțională, d.e.;
SR - cantitatea totală a mâinii drepte, PC-uri;
SL - cantitatea totală de puncte stabilite de dreapta stânga, PC-uri.
Semnul de coeficienți de asimetrie funcțională este interpretat după cum urmează: dacă valoarea coeficientului are valoarea pozitivă "+", aceasta indică un echilibru al soldului către activitatea emisferei stângi; Dacă coeficientul rezultat ia o valoare negativă, semnul "-" indică activitatea emisferei drepte.
Analizați rezultatul rezultat și încheiați.
Sinonime: Coaja de proiecție sau Colțul Departamentului de analizoare
Bark terțiar.
Pe un an, două curbe sunt pentru dreapta (albastru) și mâna stângă (roșu);
Se credea că cele mai înalte funcții ale creierului uman sunt efectuate de crusta de emisfere mari. În secolul trecut, sa constatat că atunci când eliminăm crusta la animale, ei pierd capacitatea de a îndeplini acte complexe de comportament datorită experiențelor de viață dobândite. Acum se stabilește că coaja nu este cel mai înalt distribuitor al tuturor funcțiilor. Mulți dintre neuronii săi fac parte din sistemele de motoare senzoriale și motorii la nivel mediu. Substratul funcțiilor mentale mai mari sunt sistemul de distribuție CNS, care include atât structuri subcortice, cât și neuronii cortexi. Rolul oricărui domeniu al cortexului depinde de organizația internă a legăturilor sale spinale, precum și de relațiile sale cu alte instituții de învățământ ale SNC. In orice caz. Într-o persoană în procesul de evoluție, a avut loc corticolasul tuturor, inclusiv funcțiile vibre vitale. Acestea. Supunerea lor la bază. Ea a devenit principalul sistem de integrare a întregului CNS. Prin urmare, în cazul morții unei părți semnificative a neuronilor cortexului la om, corpul său devine scăzut și moare ca urmare a insuficienței neuroostazei (hipotermie cerebrală). Creierul județean este alcătuit din șase straturi:
I. Stratul molecular, cel mai mare. Formate de multe dendrite ascendente ale neuronilor piramidei. Există puțini neuroni în el. Acest strat pătrunde axonii de nuclee de talamus nespecifice legate de formarea reticulară. Datorită unei astfel de structuri, stratul asigură activarea întregii coajă.
2 straturi de granule exterioare. Formează bine localizați neuroni mici având numeroase contacte sinaptice unul cu celălalt. Datorită acestui lucru, există o circulație lungă a impulsurilor nervoase. Acesta este unul dintre mecanismele de memorie.
3. strat de piramidă în aer liber. Constă din celule mici piramide. Cu ajutorul lor și celulelor din al doilea strat, formarea legăturilor inter-corticale, adică. Relațiile dintre diferitele zone ale crustei.
4. strat granular interior. Conține celule stea, care formează sinapse de axon de comutare și neuroni asociativi ai talamusului. Acest lucru vine aici toate informațiile de la receptorii periferici.
5. stratul intern de piramidă. Formate de neuroni mari piramizi, ale căror axe formează căi piramide descendente care se îndreaptă într-o măduvă alungită și spinării.
6. Stratul celulelor polimorfe. Aksones din neuronii lui merg la Talamus.
Neuroni corespondenți formează rețele neuronale, inclusiv trei componente principale:
1. Fibre aferente sau de intrare.
2. Interneconii
3. Neuroni de ieșire eferentă. Aceste componente formează mai multe niveluri de rețele neuronale.
1. Microseturi. Cel mai mic nivel. Acestea sunt sinapse separate interfonte cu structurile lor pre- și postsynaptice de SINAPS este un element funcțional complex având mecanisme interne de auto-reproducere. Neuronii de porumb au dendrite puternic ramificate. Ele sunt un număr mare de spini sub formă de bastoane de tambur. Aceste spini servesc pentru a forma sinapse de intrare. Sinapsele de plută sunt extrem de "" sunt sensibile la influențele externe. De exemplu, cu închisoarea de iritații vizuale, prin conținutul animalelor în creștere, duce la o scădere semnificativă a sinapselor în crusta vizuală. În cazul lui Dauna Sinaps, crusta este, de asemenea, mai mică decât normală. Fiecare Synaps de formare a spikeului, efectuează rolul convertorului de semnal care merge la neuron.
2. Rețele locale. Noua coajă este o structură stratificată, a căror straturi sunt formate din rețelele neuronale locale. Prin intermediul talamusului și a creierului olfactiv, impulsurile de la toți receptorii periferici pot veni. Fibrele de intrare trec prin toate straturile, formând sinapse cu neuronii lor. La rândul său, garanțiile fibrelor de intrare și interneuronii acestor straturi formează rețele locale la fiecare nivel al crustei. O astfel de structură a crustei oferă posibilitatea procesării, stocării și interacțiunii diferitelor informații. În plus, există mai multe tipuri de neuroni de ieșire în cortex. Aproape fiecare din stratul său oferă fibre de ieșire îndreptate către alte straturi sau zone îndepărtate ale coastei.
3. Coloane de plută. Elemente de intrare și ieșire cu Interneyronami formează coloane corticale verticale ale modulelor locale PAL. Ei trec prin toate straturile crustei. Diametrul lor este de 300-500 microni. Neuronii care formează aceste coloane sunt concentrate în jurul fibrelor cortalice care transportă un anumit tip de semnale. Există numeroase conexiuni inter-line în coloane. Neuronii 1-5 straturi de coloane oferă percepția și prelucrarea informațiilor primite. Neuroni 5-6 straturi formează căile de crustă eferentă. Coloanele învecinate sunt, de asemenea, asociate între ele. În acest caz, excitația unuia este însoțită de frânare adiacentă. În anumite zone ale crustei, coloanele sunt concentrate efectuând același tip de funcție. Aceste zone sunt numite câmpuri de cittoarchitectonice. În cortexul uman, există 53 de ele. Câmpurile sunt împărțite în primar, secundar, terțiar.
Primar furnizează procesarea unor informații senzoriale specifice.
Interacțiunea secundară și terțiară a semnalelor de diferite sisteme senzoriale. În special, câmpul de somatosensor primar, la care impulsurile provine de la toți receptorii de piele (tactil, temperatură, durere) este situată în zona de înfășurare a spatelui central. Cea mai mare parte a locului din Coreea este reprezentantul Biroului de buze, fețe, mâini ale mâinilor. Prin urmare, sub leziunile acestei zone, sensibilitatea secțiunilor de piele corespunzătoare se schimbă. Reprezentarea proproporeceptorilor de mușchi și tendoane, adică Coaja de motor ocupă un înțelept central anterior. Impulsurile de la proprigororeceptorii extremităților inferioare merg în partea de sus a înfășurării. De la mușchii corpului până la partea de mijloc. De la mușchii capului și gâtului la partea inferioară. Cea mai mare zonă a acestui câmp este, de asemenea, reprezentarea mușchilor buzelor, limbajului, a perii și fețelor.
Impulsurile receptorii de ochi intră în regiunea occipitală a cortexului în apropierea brazdei de spur. Deteriorarea câmpurilor primare duce la orbire corticală și la pierderea secundară și terțiară a memoriei vizuale. Zona auditivă a cortexului este situată în gyrusul temporal superior și transversal. Odată cu deteriorarea câmpurilor primare ale zonei, surzenia de plută se dezvoltă. Periferice - dificultăți în distingerea sunetelor. În partea treia din înfășurarea temporală superioară a emisferei stângi este centrul de vorbire al discursului - centrul Wernik. Cu schimbările patologice, abilitatea de a înțelege discursul este pierdută. Speech Center Center - Centrul Brock, este situat în urinerul frontal inferior al emisferei stângi. Încălcarea în această parte a scoarței conduc la pierderea capacității de a pronunța cuvinte.
Hemisphere de asimetrie funcționale.
Creierul frontal este format din două emisfere, care constau în acțiuni egale. Cu toate acestea, ei joacă un rol funcțional diferit. Pentru prima dată, diferențele dintre emisferele descrise 1863. Neuropatologul Paul Break. Am descoperit că, cu tumorile cota frontală stângă, pronunția capacității de a vorbi este pierdută. În anii '50 ai secolului XX, R. Serry și M. Gazzanig au investigat pacienții care, pentru a opri crizele epileptice, corpul corpului a fost făcut. Este nevoie de emisferele de legare a fibrelor de fibre. Abilitățile mintale la persoanele cu un creier split nu se schimbă. Dar, cu ajutorul testelor speciale sa constatat că funcțiile emisferelor sunt diferite. De exemplu, dacă subiectul este în câmpul de vedere al ochiului drept, Informațiile vizuale intră în emisfera stângă, atunci un astfel de pacient îl poate numi, descrie-l. Proprietățile sale. Citiți sau scrieți text.
Dacă subiectul se încadrează în câmpul de vedere al ochiului stâng, atunci pacientul nu-l poate numi chiar și spune despre el. Nu poate citi acest ochi. Astfel, emisfera stângă este dominantă în ceea ce privește conștiința, vorbirea, conturile, scrisorile, gândirea abstractă, mișcările arbitrare complexe. Pe de altă parte, deși emisfera potrivită nu are funcții de vorbire pronunțate, este capabil să înțeleagă și să gândească abstract într-o anumită măsură. Dar semnificativ mai mult decât stânga, are mecanisme de recunoaștere senzorială a memoriei eșantionului. Percepția muzicii este în întregime funcția emisferei drepte. Acestea. Dreptul emisfera este responsabil pentru funcțiile non-feribot, adică. Analiza imaginilor vizuale și auditive complexe, percepția spațiului, formele. Fiecare emisferă este izolată, reciclă și stochează informații. Ei au sentimente, gânduri, evaluări emoționale ale evenimentelor. Emisfera stângă procesează informațiile analitice, adică. în mod consecvent, și cel potrivit cel puțin, intuitiv. acestea. Emisfera utilizează diferite moduri de cunoaștere. Întregul sistem de educație din lume vizează dezvoltarea emisferei stângi, adică. Gândirea abstractă, nu intuitivă. În ciuda asimetriei funcționale, în norma emisferei lucrează împreună, furnizarea tuturor proceselor psihicului uman.
Plasticitate coajă.
Unele țesături păstrează capacitatea de a forma celule noi din celulele predecesoare pe tot parcursul vieții. Acestea sunt celule hepatice, enterocite de piele. Celulele nervoase nu posedă o asemenea abilitate. Cu toate acestea, ei mențin capacitatea de a forma noi procese și sinapse, adică fiecare neuron este capabil să fie deteriorat de nou. Restaurarea proceselor poate apărea în două moduri: prin formarea unui nou con de creștere și formare a garanțiilor. De obicei, creșterea noului axon este împiedicată de apariția cicatricei de glitch. Dar, în ciuda acestui fapt, noile contacte sinaptice sunt formate din garanții și un axon deteriorat. Cea mai înaltă plasticitate a neuronilor cortexului. Oricare din neuronul său este programat că atunci când este deteriorat, încearcă în mod activ să restabilească legăturile pierdute. Fiecare neuron este implicat și lupta competitivă cu ceilalți pentru formarea contactelor sinaptice. Acest lucru servește ca bază a plasticității rețelelor de cortex neurale. Sa stabilit că atunci când cerebelul este îndepărtat, căile nervoase merg la ea încep să germineze în coajă. Dacă în creierul intact pentru a transplanta zona creierului unui alt animal, atunci neuronii acestei bucăți de țesături formează numeroase contacte cu neuronii creierului de reciclare.
Plasticitatea cortexului se manifestă ca în condiții normale. De exemplu, în formarea unor noi legături intercornanțe în procesul de învățare și patologie. În special, funcțiile cortexului funcției sunt pierdute în timpul înfrângerii preluării câmpurilor sale vecine sau a unei alte emisfere. Chiar și cu deteriorarea zonelor extinse ale cortexului datorită hemoragiei, funcțiile lor încep să efectueze zonele corespunzătoare din emisfera opusă.
Elgcaroencefalografie. Valoarea sa pentru studii și clinici explorate.
electricefalografia (EEG este înregistrarea activității electrice a creierului de pe suprafața pielii capului. Pentru prima dată, EEG-ul omului înregistrat în 1929 psihiatrul german Gerger. La îndepărtarea EEG, sunt impuse electrozii Pielea, semnalele din care sunt îmbunătățite și hrănite la osciloscop și dispozitivul de scriere. În mod normal, se înregistrează următoarele tipuri de oscilații spontane:
1. A-Rhythm. Acestea sunt valuri cu o frecvență de 8-13 Hz. Se observă într-o stare de veghe, plină de pace și cu ochii închise. Dacă o persoană deschide ochii unui ritm înlocuiește ritmul. Acest fenomen se numește blocada unui ritm.
2. B-ritm, frecvența sa de la 14 la 30 Hz. Se observă cu o stare activă a creierului și este citită ca intensitatea lucrărilor mentale crește.
3. (Gama) - ritm. Oscilații cu o frecvență de 4-8 Hz. Înregistrat în timpul adormirii adormite.
somnul de suprafață și anestezia superficială.
4. (Sigma) - ritm. Frecvență 0,5-3,5 Hz. Observate cu somn și anestezie profundă.
Cu cât este mai mică frecvența ritmurilor EEG, cu atât amplitudinea lor. În plus față de aceste ritmuri majore, sunt înregistrate alte fenomene EEG. De exemplu, ca dormit este aprofundat, apar axele somnoroase. Aceasta este o creștere periodică a frecvenței și amplitudinii ritmului Theta. Când așteptăm echipa la acțiune, apare un val negativ de așteptare etc.
Experimentul EEG este utilizat pentru a determina nivelul activității creierului și în clinica pentru diagnosticarea epilepsiei (în special a formelor ascunse), precum și pentru a identifica moartea creierului (coaja trăiește 3-5 minute, neuronii stem 7 -10, inima 90. Rinichii 150).
Funcțiile măduvei spinării
În substanța albă a măduvei spinării, este localizată adiacentă substanței gri dintre coarnele din față și spate, formarea reticulară. Această formare este formată din clustere de celule nervoase care au numeroase conexiuni între ele. R. formarea eticulară Oferă activitatea altor neuroni ai măduvei spinării datorită proprietății de automatizare (vezi mai jos).
Reflexe vegetative (Vasomotor, transpirație, urinară, defect) se datorează prezenței în măduva spinării veganilor sistemului nervos vegetativ (vezi mai jos).
Explorați funcțiile
Acestea sunt efectuate conform legii lui Bella Majandi: informațiile aferente intră în măduva spinării prin rădăcinile din spate, impulsurile eferente sunt transmise prin rădăcinile din față.
Creșterea căilor (sensibile) Măduva spinării este situată în piloții din spate Alb. Substanțe și informații de transport din lumea exterioară și mediul interior al organismului:
1) de la receptorii pielii (durere, temperatură, atingeri, presiune, vibrații);
2) de la proprigororeceptori (spindlere musculare, receptorii de tendre din Golgi, vasele și cochilii articulate);
3) de la receptorii organelor interne - visceororeceptoare (mecanice și chimioreceptoare).
Căi descendente (motor) Situat în stâlpii din față și transmite impulsuri la mușchii scheleticii despre mișcările arbitrare (conștiente), efectele tonice asupra mușchilor, impulsurile care asigură menținerea posturii și echilibrului. În căile descendente sunt, de asemenea, transmise efecte vegetative (pe organele interne).
Funcții dirijoare similare și în alte structuri stem (creier obținut, creier mediu și pod): pe grupul din spate al fibrelor albe sunt căi de conducere aferente, pe front-eferente.
Funcțiile creierului alungit
De bază funcția Pyramid - Conducerea semnalelor despre mișcările arbitrare.
Funcțiile de bază ale măslinelor sunt legate de menținerea echilibrului.
În creierul alungit sunt situate kernel-uri ale nervilor cranieni VIII-XII,prin urmare, creierul alungit efectuează reflexe protectoare (tuse, strănut, vărsături, lacrimi, închidere, o îngustare a elevului) (vezi).
Brainul alungit exercită funcții senzoriale: recepția sensibilității pielii față, analiza primară a gustului. Există semnale de la chimioreceptori și baroreceptori ai navelor, organelor interne interioare și a vaselor vestibulare la curajos. Influența acestor structuri determină funcționarea la nivelul creierului alungit centrele respiratorii, inimii și vasculare. Structura formării reticulare îndeplinește, de asemenea, funcțiile de reglare tonul musculaturii scheletului.
Funcții conductive - vezi măduva spinării.
Structurile creierului din spate
Creierul din spate include un pod și cerebelum.
Pod facial (Pereche VII) și pRE-DOOR-SENLAT (Cuplu VIII) Nervii.
Responsabil pentru răspunsul fiziologic al tensiunii și anxietății, participă la mecanismele de somn.Mulți dintre neuronii lui noradrenergic.
Funcții de pod:
· Conductoare (predomină);
· Oferă menținerea poziției și conservarea echilibrului corpului în spațiu în momentul schimbării vitezei mișcării;
· Oferă tonul mușchilor cervicali;
· Conține centre vegetative de reglare respiratorie (centru de pneumografie), ritm cardiac, activitate a tractului gastro-intestinal.
· Reglează mestecarea și înghițirea (a se vedea Reflexe complexe ale creierului);
· Joacă un rol important în activarea coaja de emisfere mari (inclusiv într-o stare de anxietate);
· Limitează intrările senzoriale ale impulsurilor nervoase la emisfere mari în timpul somnului.
Cerebellum.
Funcțiile cerebelului sunt în principal legate de organizarea actelor motorii și reglarea funcțiilor vegetative. De la cortexul motorului și cerealele bazale din Cerebelum vine informații despre mișcarea planificată, precum și aferența din sistemul somatosensorial. Cerebelum oferă coordonarea reciprocă a mișcărilor, precum și corectarea traficului efectuată(necesar, deoarece punerea în aplicare a unui act motor, părțile mobile ale corpului experimentează efectul forțelor inerțiale, care perturbă netezimea și acuratețea mișcării).
Funcții Cerebellum:
· Salvarea punctelor și echilibrului corpului;
· Coordonarea mișcărilor vizate;
· Construirea mișcărilor balistice rapide;
· Reglementarea tonului muscular;
· Reglementarea funcțiilor vegetative (bătăi de inimă, ton de nave, motociclete intestinale etc.);
· Explorer.
Funcțiile creierului de mijloc
În creierul mijlociu, ele alocă o localizare dorsală acoperiş și mersul ventilului picioarele creierului.
formarea reticulară, nucleii oxtomotory și bloc Nervii de carduri (aburul III-IV).
Acoperișul creierului mijlociu constă din patru elevi ( quinoa) - Holmikov, având o vedere a emisferei.
Picioarele creierului Două roți groase, longitudinale care merg la emisferele drepte și stângi ale unui creier mare. În picioarele mai groase ale trecerii creierului miezurile substanței neagră. În anvelopele se află kerneluri extrapiramine (nuclee roșii, substanță neagră si etc.).
Crăciunul nervilor Chereful (III-V)și formarea reticularăparticipați la vânzări reflexe de creier complexe.
Substanță neagră - una dintre secțiunile creierului care produc dopamină. În plus, substanță neagrăefectuează o serie de funcții esențiale: reglementarea tonului muscular, în special în timpul somnului, furnizarea de homeostază este inclusă în sistemul anti-alunecare și formare a somnului organismului.
Reacții toniceÎmpreună cu poziția măduvei spinării, redistribuirea tonului diferitelor grupări musculare la schimbarea poziției corpului sau a părților sale individuale (de exemplu, capul) în spațiu. Ele sunt împărțite în două grupe: statice și sebrinetice. Reacții statice Se întâmplă atunci când poziția corpului este schimbată, care nu este legată de mișcarea sa în spațiu (adică reflexele poziției). Reacții Stocietice se manifestă în redistribuirea tonului muscular scheletic, care asigură conservarea echilibrului corpului uman la accelerațiile unghiulare și liniare ale mișcării active sau pasive în spațiu
Creierul intermediar
Creierul intermediar Aceasta este cea mai mare diviziune a creierului, a cărei cavitate este III Stomac.. Creierul intermediar este situat sub corp de porumb și arc Creierul, cea mai mare parte este înconjurată de emisferele creierului final. Intermediarul creierului include umflături vizuale (talamus), subburo (hipotalamus), partea de propagandă (epicitulamus) și regiunea de custodie (metatalamus). Creierul intermediar include, de asemenea, două glande de secreție internă - pituitar. și epiphysis. (corpul cishemoid).
Talamus.
Talamus (lovituri vizuale)reprezintă acumularea de substanțe gri, au o formă ovoidă, conectată interstalama Spike.. Celulele nervoase sunt grupate într-un număr mare de nuclee (până la 120). Funcțional a kernelului de talamus este împărțit în specific, nespecifică, asociativ și motor.
Nuclee specifice asociate cu anumite zone de cortex sensibile - auditive, vizuale etc. (toate, cu excepția Olfactory). Aici există o convergență a semnalelor aferente cu suprimarea biologică nesemnificativă. Nuclee nespecifice Talamusul este asociat cu multe secțiuni ale coastei și, împreună cu structurile formării reticulare, iau parte la formarea unor influențe ascendente de activare. Nucleul asociativ Formată multipolară, ale cărei axonii merg la straturile de zone asociative și parțial de proiecție. Kernel-urile asociative sunt implicate în procese integrative mai mari (corvoare multisensor, etc.), dar funcțiile lor nu sunt încă suficient de studiate. LA motor nuclear Talamusul aparține miezului ventral, care are o intrare din cerebelul și ganglionii bazali și, în același timp, oferă proiecție în zona motorului de emisfere mari. Acest kernel este inclus în sistemul de reglare a mișcării.
Hipotalamus
Hipotalamusformează pereții și fundul celui de-al treilea ventricul, se blochează de pe un picior subțirepituitar. . În hipotalamus iese în evidență trei zone de nuclee de cluster: față, mediu (medial) și spate. În zona din față Hydallamus este situat supratoptic.și kerneluri paraventriculare. În celulele neurosecretoriale ale acestor nuclee, sunt produse hormoni, care intră în miza din spate a hipofizei (neurohypophysis). În zona mijlocie (medială) Neuroni situați în cazul în care sunt produse neurogononii liberina. și statine, respectiv, activarea sau deprimarea funcționării lobului din față al hipofizei adenogipofiz). La nucleare zona din spate Acestea sunt celule mari împrăștiate, precum și kernel-uri. corpul de vară.
Hipotalamusul este structura CNS, care transportă complicată integrarea funcțiilor diferitelor organe internela activitatea integrală a corpului. Modifică activitățile sistemelor viscerale cardiovasculare, respiratorii și alte sisteme viscerale cu schimbări în mediul extern sau intern (modificări ale condițiilor meteorologice, exerciții fizice, infecții etc. care amenință homeostazia). În funcție de executiv funcții vegetative Există două zone în hipotalamus. Prima zonă dinamogenic., ocupând partea mijlocie și din spate a hipotalamusului. Când este încântat, se observă "reacțiile motorii": extinderea elevului, creșterea bătăilor inimii, creșterea tensiunii arteriale, activarea respiratorie, creșterea excitabilității motorului, adică manifestări ale influențelor simpaticesistemul nervos vegetativ. A doua zonă este tropogenExcitarea sa se manifestă într-o îngustare a elevului, scăderea tensiunii arteriale, demisia respirației, vărsăturilor, defectării, urinării, salivei, adică Simptomele caracteristice influențele sistemului nervos parasympatic.
În hipotalamus sunt situate centrele motivaționale: foame, saturație, sete, precum și centre de sex și agresiv-defensive. Având obținerea unor fluxuri aferente de excitație de la interiorsteceptori (osimorceptori, chimioreceptori, termistori etc.) și integrarea acestora cu influențe umorale asupra celulelor nervoase ale hipotalamusului, aceste centre formează stările motivaționale corespunzătoare ale corpului.
SISTEMUL LYMBIC.
SISTEMUL LYMBIC. (Sinonim: un complex limbă, creier visceral) - un complex al structurilor creierului mijlociu, intermediar și finit implicat în organizarea reacțiilor viscerale, motivaționale și emoționale ale corpului. Formularul sistemului limbic: bulbul olfactiv; tractul olfactiv; triunghi olfactiv; Substanța forțată frontală; șoc de talie; paragippocampal convoluție; hipocampus.; corpul în formă de migdale; hipotalamus; corp superior; formarea reticulară mezencefal.
Sistemul limbic este redat influența modulativă pe coaja unui creier mare și a structurilor subcorticale, stabilind necesară o formare reticulară nivelul activității lor(Ascendent: Coma → Somnul adânc → Somnul de suprafață (Dund) → Să trezește calm → Vegherea activă → Starea excitată → Afecțiune). Sistemul limbic controlează emoțiile, ciclul de somn-trezire, comportamentul sexual, precum și procesele de învățare și memorare. La primirea informațiilor despre mass-media exterioară și interioară a corpului, sistemul limbic lansează reacții emoționale vegetative și somatice (creșterea bătăilor inimii și respirația, o creștere a tensiunii arteriale și a transpirației, tensiunii musculare). Formațiile de liimitate se referă la centre integrative mai mari reglementarea caracteristicilor vegetative ale corpului. Din ele, impulsurile de excitație sunt trimise la centrele de hipotalamus vegetativ și prin intermediul acestuia la hipofiza și stem și nucleele spinoase ale sistemului nervos autonom. Datorită legăturilor sale cu ganglionii bazali, departamentele din față ale talamusului și formarea reticulară, formațiunile limbice pot afecta tonul mușchilor scheletici.
O caracteristică a activității sistemului limbic este că există legături bilaterale simple între structurile sale și căile complexe care formează o varietate de cercuri închise ( cercul Peypes.). O astfel de organizație creează condiții pentru circulația pe termen lung a aceleiași excitație în sistem și, prin urmare, menținerea unui singur stat și impunând acest stat la alte sisteme ale creierului ( excitație reverb.). Aceasta determină nu numai activarea tonică a cortexului creierului mare, ci și puterea și severitatea stărilor emoționale ale corpului; Legate de memoria și procesele de instruire și memorie pe termen scurt, reglementează forme agresive defensive, alimente și sexuale de comportament.
Nucleii bazali.
În substanță albă, emisferele creierului, mai aproape de baza sa, substanță cenușie durtă care formează kerneluri subcortice sau bazale: corpul dungiconstând din coadă Nucleii Ichchevice (include coajă, minge palidă laterală și palidă), garduri, corp în formă de migdale.
Bazale otrăvit central la structuri sisteme de mișcări arbitrare. (motocanele motorului). Odată cu participarea nucleelor \u200b\u200bbazale, sinergismul tuturor elementelor unor astfel de acte complexe de motor, cum ar fi mersul pe jos, alergând, alpinism; Se realizează netezirea mișcărilor și instalarea posturii inițiale pentru implementarea acestora. Kernel-urile bazale coordonează tonul și activitatea motorului de fază a mușchilor. Activitatea lor este legată de implementarea mișcărilor lente, cum ar fi mersul pe jos, depășind un obstacol, inventarea firelor în ac.
Miezurile bazale participă nu numai în reglementarea activităților motorii, ci și în analiza fluxurilor aferente, în reglementarea unui număr de funcții vegetative, în implementarea unor forme complexe de comportament congenital, în mecanismele de memorie pe termen scurt , precum și în reglarea ciclului de somn-Wake.
Funcțiile cortexului de emisfere mari
Cel mai înalt departament al CNS este mare hemispieie mare. Diferitele zone ale cortexului creierului mare au câmpuri diferite care sunt determinate de natura și numărul de neuroni, grosimea straturilor etc. Prezența câmpurilor structurale diferite implică diferit scopul lor funcțional.
Luând în considerare caracteristicile funcționale ale câmpului Noua coajă, împărțită în primar, secundar și terţiar sau asociativ. Câmpurile primare și secundare combină departamentele de coajă asociate cu funcționarea anumitor sisteme senzoriale.
1) Câmpurile primare (proiecție) primesc și procesează informații din orice sistem senzorial. Aici este efectuată analiza primară Informații senzoriale într-o modalitate (de exemplu, pentru culoarea vizuală, iluminare, formă). Modalitate - tipul de senzații senzoriale - auditiv, vizual, olfactiv, etc.
Câmpurile principale senzoriale și motoare sunt strict localizate. Următoarele sunt unele dintre ele.
În nucleul de înfășurare post-centrală și felii parietale superioare, celulele nervoase care formează proprioceptive și sensibilitate generală (Temperatură, durere și tactilă). Analizor motor Core. Situat în zona motorului coastei la care întinderea precentrală și tăierea paracanitală a suprafeței emisferei mediale. Mărimea și amplasarea zonelor de proiecție ale diferitelor organe din crusta sensibilă la aragaz și motor depind de valoarea lor funcțională.
În adâncurile canelurii laterale, pe suprafața de înfășurare temporală superioară cu fața insulei, este kernel-ul analizorului auditiv. În cortexul de înfășurare temporală medie miezul analizorului vestibular.
Miezul analizorului vizual Acesta este situat pe suprafața mediană a acțiunii occipitale, pe ambele părți ale brazdei spur.
SPECIALE CENTRELOR Situat în mâna dreaptă în emisfera stângă, iar partea stângă este în dreapta. Analizor motor de vorbire Core (Rech recreak) este situată în departamentele din spate ale înfășurării frontale inferioare ( centrul Brock.). Kernelul analizorului auditiv al discursului oral (Percepția vitezei) este strâns legată de centrul auditiv corticular și se află în secțiunile din spate ale înfășurării temporale superioare, pe suprafața sa cu fața spre brazda laterală ( zone Broom.). Lângă miezul analizorului vizual este nucleul analizorului vizual al discursului scris.
Cork departamente gust și olfatoriu Analizorii se află pe suprafața inferioară a acțiunii temporale, în înfășurarea skatei de mare și a cârligului pe suprafața inferioară a acțiunii temporale.
2) Câmpurile secundare sunt situate deasupra primar și ocupă o zonă mare. În plus față de fibrele sensibile, din centrele motivaționale și emoționale, structurile de memorie etc. vin la ele. Pentru ei este caracteristică identificare Imagini senzoriale În cadrul unei modalități (de exemplu, recunoașterea subiectului este un cui, șurub, tija, diblu, călcâi, ciupercă, mamelon, ac). Deteriorarea domeniilor secundare pot duce la agnozioane senzoriale (încălcări ale proceselor de recunoaștere): gust vizual, auditiv, olfactiv, gust, precum și amphiia senzorială (încălcarea recunoașterii vorbirii).
3) Câmpurile terțiare sau asociative ocupă mai mult de 50% din întreaga suprafață de emisfere și sunt cei mai tineri (în termeni evolutivi). Câmpurile terțiare au o legătură strânsă cu nucleele asociative ale Talamusului. Zonele asociative oferă contacte între zonele de proiecție ale analizoarelor individuale și integrează activitățile acestora. Acestea participă la procesarea multi-senzor a informațiilor, formând răspunsuri și implementarea unor forme complexe de comportament. În plus, există și alte tipuri de convergență: senzoriale-biologice (se manifestă în ajutorul neuronilor individuali ai cortexului creierului mare al excitațiilor aferente oricăror modalități senzoriale și excitații motivaționale asociate cu diferite stări biologice ale corpului (durere, foame, etc.), multibiologice și eferente aferente. Principalele zone asociative sunt darmer-Zatilochny. (în principal funcția de percepție) și frontal (Organizarea și controlul comportamentului, în principal motor, reacții). Departamentul frontal din față este substrat morfologic al activității mentale (conștiință, gândire, învățare, memorie, emoții).
1. Barurarea mare de emisferă efectuează funcția celei mai mari analize a semnalelor de la toți receptorii corpului și cea mai mare sinteză a răspunsurilor la actul fezabil din punct de vedere biologic.
2. Big emisferele mari reprezintă cea mai mare coordonare a organelor de activitate a activității reflexe. Ea este capabilă să renunțe la rândul său, încetinește. Coordonează activitatea departamentelor subiacente, etajele SNC.
3. Coaja de emisfere mari, ca cel mai mare corp de coordonare a activității reflexe, formează reacții recomandate din punct de vedere biologic care asigură adaptarea corpului în mediul extern, reacții care echilibrează organismul cu mediul extern.
4. În cea mai înaltă etapă a dezvoltării sale a sistemului nervos central, coaja mare cu inima dobândește o altă caracteristică, devine un corp mental. Pe baza proceselor fiziologice există senzații și percepții în el, apare gândirea. Cortexul creierului este un corp de gândire. Creierul uman, cea mai înaltă graniță a emisferelor mari, oferă posibilitatea vieții sociale, oferă posibilitatea de comunicare, cunoașterea lumii înconjurătoare, cunoașterea naturii.
Anatomia și histologia crustei
Big emisfere mari - cel mai perfect aparat CNS. Ea și-a urcat pe nume, deoarece acoperă creierul din toate părțile, pe măsură ce coaja de copac îi înconjoară butoiul. Este ridicată de o mulțime de brazdă și sorrus. De mai sus, este acoperită cu un strat de neuroni, grosimea căreia variază în intervalul de 2-4 mm, constituindu-se pe o medie de 2,5 mm. Există aproximativ 49 de miliarde de celule în crustă, adică. 14/15 Toți neuronii. (Începând de la 20 de ani, aproximativ 100 mii de neuroni cortexi mor în fiecare zi). Partea principală a scoarței constă în substanță albă. Substanța albă a creierului frontal este formată din axonii acestor celule, precum și axonii de căi diferite în creștere. Ca și în fiecare centru nervos, există neuroni senzoriali în cortex care percep informații de la aducerea căilor, neuroni eferenți care trimit comenzi pe căile descendente și inservează sau neuroni asociativi care constituie cea mai mare parte a masei. Datorită proceselor de neuroni asociativi, coaja este combinată într-un singur întreg: excitarea a apărut într-un singur loc poate acoperi întreaga coajă.
În funcție de filogeneză, în conformitate cu istoricul dezvoltării coaja de emisfere mari, se disting 3 părți.
1. Coarta antică - arhocriticism. Coaja antică include becurile olfactive (fibrele aferente vin aici din epitelia olfactivă a mucoasei nazale), se află tractele olfactive (situate pe suprafața de jos a acțiunii frontale) și tuberculii olfactivi (se află centrele secundare olfactive).
2. Bark vechi - Paleokortex. Coaja veche se dovedește a fi certată la Ulusulină, Hipocampus și migdale. Toate aceste formațiuni fac parte din sistemul limbic, care este cel mai înalt departament al sistemului nervos vegetativ.
3. Noua Cora - Neokortex. Compoziția noii coajă include toate celelalte zone ale cortexului de emisfere mari: acțiuni frontale, temporale, occipitale, parietale.
În procesul de filogeneză, noua coajă apare mai întâi la mamifere și ajunge la o dezvoltare mai mare la om, adică este cea mai tânără structură nervoasă, iar la om exercită cea mai mare reglementare a funcțiilor corpului și a proceselor psiho-fiziologice care oferă diferite forme de comportament.
Citoararchitectonica crustei (Localizarea și interreclația neuronilor în crustă). Dacă coaja antică are 3 straturi, atunci coaja nouă are o construcție de 6 straturi.
1. Stratul superficial este molecular. În acest strat există foarte puține celule nervoase, dar multe fibre de ramificare ale celulelor subiacente care formează o rețea groasă de plexuși.
2. Cel de-al doilea strat este o granulă exterioară, prezentată în celule de bază și celule piramide parțial mici. Fibrele celulelor din cel de-al doilea strat sunt amplasate în principal de-a lungul suprafeței cortexului, formând conexiuni cortico-corticale.
3.Tresează stratul - stratul de piramidă exterioară constă în principal din celulele de piramidă mijlocie. Axele acestor celule, cum ar fi stratul de celule granulare II, formează conexiuni asociative cortic și corticale.
4 strat interior granular în funcție de natura celulelor (celulele stea) și fibrele lor sunt similare cu stratul exterior de cereale. În acest strat, terminațiile sinaptice ale fibrelor aferente provenind din neuronii nucleelor \u200b\u200bspecifice ale talamusului; Cea mai mare densitate capilară este observată aici.
5. Stratul de piramidă internă sau stratul de celule BETZ. Acest strat constă în principal din celule de piramide medii și mari. Dar, în acest strat într-un prezentator este o celulă piramidală mare, gigant, celule ale BETZ. Dendritele lungi ale acestor celule sunt împinse și ajung la stratul de suprafață - acestea sunt așa-numitele dendrite apicale. Axonii celulelor BETZ merg la diferite nuclee ale capului și a căilor motociclete colico-spinării colaboratorului spinal și cortic. Cele mai lungi axe fac parte din calea piramidală și ajung la segmentele inferioare ale măduvei spinării, terminând celulele introduse și pe mecanismele A-Motor-Motor al măduvei spinării.
6. Stratul de celule polimorfe se formează în mod predominant celule în formă de coloană a căror axonii formează căi cortico-talantice.
Impulsurile aferente de intrare intră în coaja de mai jos, se ridică la celulele ⅲ - ⅴ straturile cortexului, există percepția și prelucrarea semnalelor care intră în coajă.
Principalele obligațiuni eferente ale cortexului de emisfere mari sunt escortarea căilor eferente care sunt în principal în straturile V-VI.
Diviziunea mai detaliată a cortexului în diverse domenii a fost efectuată pe baza semnelor citoarhitectonice K. Brodman (1909), care a alocat 52 de câmpuri; Multe dintre ele se caracterizează prin caracteristici funcționale și neurochimice.
Datele histologice arată că lanțurile neuronale elementare implicate în procesarea informațiilor sunt localizate perpendiculare pe suprafața crustei. În nucleul creierului există asociații funcționale de neuroni situați într-un cilindtrix cu un diametru de 0,5-1,0 mm. Aceste sindicate au fost numite difuzoare neurale . Ele se găsesc în cortexul motorului, în diferite zone ale cortexului senzorial. Coloanele neuronale învecinate pot interacționa între ele.
Astfel, diferite zone ale coajei noi au o structură clară și stereotipică.
Dar, în ciuda rușinii organizației neurale a întregului coajă, diferite departamente de coajă diferă unul de celălalt. Distinsa concluziile în cantitatea și dimensiunea neuronilor, în timpul fibrelor, ramificarea axonilor și a dendritelor. Aceste diferențe se datorează funcției inegale a diferitelor zone ale crustei. Fiecare secțiune, zona cortexului efectuează o funcție definită, există o specializare funcțională a diferitelor zone ale crustei.