Om de betekenis van de structurele kenmerken van de cel niet alleen te begrijpen, maar ook het belangrijkste, om de functionele secties van zijn individuele componenten en de gehele cel in het algemeen te begrijpen, om de studie van de celmorfologie te combineren met de hoofdbiochemische en genetische kenmerken van zijn inrichting en werk om de cel precies de posities van de moderne celbiologie te bestuderen, het is noodzakelijk om onmiddellijk de belangrijkste moleculaire biologische patronen te herinneren, wederom, kortweg verwijzen naar de inhoud van het centrale dogma van de moleculaire biologie.
Cel als zodanig voert een verscheidenheid aan functies uit. Zoals we al hebben gesproken, zijn sommige van hen over het algemeen cel, sommige zijn speciaal, kenmerkend voor speciale celtypen. De belangrijkste werkmechanismen voor het uitvoeren van deze functies zijn eiwitten of hun complexen met andere biologische macromoleculen, zoals nucleïnezuren, lipiden en polysacchariden. Het is bijvoorbeeld bekend dat transportprocessen in verschillende stoffen, beginnend met ionen en eindigend met macromoleculen, worden bepaald door de werking van speciale eiwitten of lipoproteïnecomplexen die deel uitmaken van plasma en andere celmembranen. Bijna alle processen van synthese, verval, herstructurering van verschillende eiwitten, nucleïnezuren, lipiden, worden koolhydraten optreden als gevolg van activiteit die specifiek is voor elke individuele reactie van enzymeiwitten. Synthese van individuele biologische monomeren, nucleotiden, aminozuren, vetzuren, suikers en andere verbindingen worden ook uitgevoerd door een enorm aantal specifieke enzymen - eiwitten. De vermindering die leidt tot celmobiliteit of aan de beweging van stoffen en structuren in de cellen wordt ook uitgevoerd door speciale contractiele eiwitten. Veel celreacties in reactie op de impact van externe factoren (virussen, hormonen, buitenlandse eiwitten, enz.) Begin met de interactie van deze factoren met speciale receptorcellen.
Eiwitten zijn de belangrijkste componenten van bijna alle cellulaire structuren. Veel chemische reacties in de cel worden bepaald door de set enzymen, die elk een of meer individuele reacties leiden. De structuur van elk individueel eiwit wordt strikt gespecificeerd, dat wordt uitgedrukt in de specificiteit van hun primaire structuur - in de aminozuursequentie langs het polypeptide, eiwitketen. Bovendien is de specificiteit van deze aminozuursequentie onmiskenbaar herhaald in alle moleculen van dit cellulaire eiwit.
Een dergelijke correctheid bij het reproduceren van de ondubbelzinnige sequentie van aminozuren in de eiwitketen wordt bepaald door de structuur van het DNA van het gengedeelte, dat uiteindelijk verantwoordelijk is voor de structuur en synthese van dit eiwit. Deze ideeën dienen als de belangrijkste postulate van moleculaire biologie, het "dogma". Informatie over het toekomstige eiwitmolecuul wordt verzonden naar de locatie van zijn synthese (in ribosomen) door een tussenpersoon - informatie-RNA (IRNA), waarvan de nucleotidesamenstelling de samenstelling en volgorde van het nucleotide van het DNA-gen weerspiegelt. Een polypeptideketen is gebouwd in het ribosoom, de sequentie van aminozuren waarin wordt bepaald door de sequentie van nucleotiden in de IRNA, de reeks van hun triplets. Aldus benadrukt het centrale dogma van de moleculaire biologie de unidirectionaliteit van informatieoverdracht: alleen van DNA naar het eiwit met behulp van een tussenliggende link - IRNA (DNA → IRNA → eiwit). Voor sommige RNA-bevattende virussen kan de informatietransmissieketen volgens het RNA-schema → Irna → eiwit gaan. Het verandert de essentie van de zaak niet, omdat de bepalende link hier ook nucleïnezuur is. Omgekeerde manieren van vastberadenheid van eiwit tot nucleïnezuur, naar DNA of RNA zijn onbekend.
Om verder te gaan naar de studie van celstructuren die geassocieerd zijn met alle stadia van eiwitsynthese, moeten we in het kort verblijven op de belangrijkste processen en componenten die dit fenomeen bepalen.
Momenteel kan op basis van moderne ideeën over de biosynthese van eiwitten, de volgende algemene hoofdschema van dit complexe en meertrapsproces worden gegeven (fig. 16).
Het belangrijkste, "team", de rol bij het bepalen van de specifieke structuur van eiwitten behoort tot deoxyribonucleïnezuur - DNA. Het DNA-molecuul is een extreem lange lineaire structuur bestaande uit twee intercloine-polymeerketens. Samengestelde elementen - Monomeren - deze kettingen zijn vier variëteiten van deoxyribonucleotiden, afwisseling of sequentie waarvan uniek is langs de keten en specifiek voor elk DNA-molecuul en elk van zijn secties. Verschillende voldoende lange delen van DNA-moleculen zijn verantwoordelijk voor de synthese van verschillende eiwitten. Een DNA-molecuul kan dus de synthese bepalen van een groot aantal functioneel en chemisch verschillende cel-eiwitten. Voor de synthese van elk type eiwitten is slechts een bepaald deel van het DNA-molecuul verantwoordelijk. Een dergelijk deel van het DNA-molecuul dat is geassocieerd met de synthese van een ander eiwit in de cel wordt vaak aangeduid met de term "CYSTRON". Momenteel wordt het concept van Cyastron beschouwd als een equivalent concept van gen. In de unieke genstructuur, in een bepaalde sequentiële opstelling van zijn nucleotiden langs de keten, wordt alle informatie over de structuur van één overeenkomstig eiwit afgesloten.
Uit de algemene regeling van eiwitsynthese wordt het gezien (zie figuur 16) dat het initiële item waaruit de informatiestroom voor de biosynthese van eiwitten in de cel begint, DNA is. Daarom is het het DNA dat de eerste verslag van informatie bevat die moet worden gehandhaafd en gereproduceerd van de cel naar de cel, van generatie tot generatie.
Kort aanraken van de vraag over de plaats van opslag van genetische informatie, d.w.z. Op de lokalisatie van DNA in de cel kunnen we het volgende zeggen. Het is al lang bekend dat het DNA, in tegenstelling tot alle andere componenten van het Whiteoxithating-apparaten, een speciale, zeer beperkte lokalisatie heeft: de locatie in de cellen van hogere (eukaryote) organismen zal de celkern zijn. In de lagere (prokaryote) organismen die geen ingerichte celkern hebben, wordt DNA ook opgemerkt van de rest van het protoplasma in de vorm van een of meer compacte nucleotidevormingen. In volledige naleving hiermee is de eukaryote kernel of de prokaryote nucleïde al lang beschouwd als een aanbieder van genen als een unieke cellulaire organoïde, die de implementatie van erfelijke tekenen van organismen en hun transfer tot generaties controleert.
Het belangrijkste beginsel dat ten grondslag ligt aan de macromoleculaire structuur van DNA is het zogenaamde complementaire principe (fig. 17). Zoals reeds vermeld, bestaat het DNA-molecuul uit twee interclowed-ketens. Deze ketens zijn met elkaar verbonden door de interactie van hun tegenovergestelde nucleotiden. Tegelijkertijd is volgens structurele redenen het bestaan \u200b\u200bvan een dergelijke kruik-in-kamerstructuur alleen mogelijk als de tegenovergestelde nucleotiden van beide ketens sterisch complementair zullen zijn, d.w.z. zal je ruimtelijke structuur zijn om elkaar aan te vullen. Een dergelijke complementaire - complementaire - paren nucleotiden zijn stoom A-T (Adenin-Timin) en een paar van Mr. (Guanin-Cytosin).
Daarom, volgens dit principe van complementariteit, als we een bepaalde sequentie van vier variëteiten van nucleotiden in één keten hebben, dan in de tweede keten, zal de sequentie van nucleotiden ondubbelzinnig worden bepaald, zodat elk en de eerste keten overeenkomt met de Tweede ketting, elke eerste keten - en in de tweede keten, is elke G van de eerste keten C in de tweede keten en elke C eerste keten - R in de tweede keten.
Het gespecificeerde structurele principe dat ten grondslag ligt aan de schuimstructuur van het DNA-molecuul maakt het gemakkelijk om de exacte reproductie van de bronstructuur, d.w.z. te begrijpen Exacte weergave van informatie die is vastgelegd in de molecuulcircuits in de vorm van een bepaalde sequentie van vier variëteiten van nucleotiden. Inderdaad, de synthese van nieuwe DNA-moleculen in de cel gebeurt alleen op basis van de reeds bestaande DNA-moleculen. Tegelijkertijd beginnen de twee ketens van het oorspronkelijke DNA-molecuul zich te verspreiden van een van de uiteinden, en op elk van de gescheiden single-secties begint te worden verzameld uit de tweede keten die aanwezig is in het medium vrije nucleotiden bij nauwkeurig naleving met het principe van complementariteit. Het proces van discrepantie tussen twee ketens van het oorspronkelijke DNA-molecuul gaat door, en dienovereenkomstig worden beide ketens aangevuld met aanvullende circuits. Dientengevolge (zoals te zien in Fig. 17), in plaats van één, ontstaan \u200b\u200btwee DNA-moleculen, precies identiek aan het origineel. In elk van het resulterende "dochter" DNA-molecuul is één ketting volledig te wijten aan het origineel, en de andere wordt wonderlijk gesynthetiseerd.
Het is noodzakelijk om te benadrukken dat het potentiële vermogen om nauwkeurig te reproduceren wordt gelegd in de kleinste complementaire DNA-complementaire structuur als zodanig, en de ontdekking hiervan is absoluut een van de belangrijkste prestaties van de biologie.
Het probleem van reproductie (reductioneel) DNA is echter niet uitgeput door de statuut van het potentiële vermogen van de structuur om zijn nucleotidesequentie nauwkeurig te reproduceren. Het feit is dat het DNA zelf helemaal geen zelfreproductiemolecuul is. Om het syntheseproces te implementeren - de reproductie van DNA volgens de hierboven beschreven regeling - de activiteit van een speciaal enzymatisch complex, dat de naam is van het DNA-polymerase noodzakelijk. Het is dit enzym dat het DNA-molecuul consequent van het ene uiteinde aan een ander proces van discrepanties van twee ketens met gelijktijdige polymerisatie op hen, gratis nucleotiden volgens het complementaire principe. Aldus stelt DNA, zoals een matrix, alleen de volgorde van het nucleotide in de synthetiserende ketens, en het proces zelf leidt eiwit. Het werk van het enzym tijdens de vermindering van DNA is een van de meest interessante problemen vandaag. Het is waarschijnlijk een DNA-polymerase omdat het actief langs het twintig DNA-molecuul van het ene uiteinde naar de andere moet kruipen, waardoor een split verminderde "staart" achter. De fysieke principes van dit werk van dit eiwit zijn nog niet duidelijk.
DNA en zijn individuele functionele secties die informatie over de structuur van eiwitten dragen, directe deelname zelf in het proces van het maken van eiwitmoleculen worden niet geaccepteerd. De eerste fase op weg naar de uitvoering van deze informatie die is opgenomen in DNA-kettingen is het zogenaamde transcriptieproces of "herschrijven". In dit proces, op dezelfde DNA-circuit, zoals op de matrix, treedt de synthese van chemisch gerelateerd polymeer - ribonucleïnezuur (RNA) voor. Het RNA-molecuul is één ketting, waarvan de monomeren vier variëteiten van ribonucleotiden zijn, die worden beschouwd als een kleine wijziging van de vier variëteiten van DNA Deoxyribonucleotiden. De sequentie van de vier variëteiten van ribonucleotiden in het resulterende RNA-circuit herhaalt exact de sequentie van de overeenkomstige deoxyribonucleotiden van een van de twee DNA-circuits. Op deze manier wordt de nucleotidesequentie van genen gekopieerd als RNA-moleculen, d.w.z. De informatie die is opgenomen in de structuur van dit gen is volledig herschreven op RNA. Een groot, theoretisch onbeperkt aantal van dergelijke "kopieën" - RNA-moleculen kan uit elk gen worden verwijderd. Deze moleculen, herschreven in vele exemplaren van beide "kopieën" van genen en, om dezelfde informatie als genen te dragen, divergeren via de kooi. Ze zijn al direct bij de cellen van de cellen opgenomen en accepteren "Persoonlijke" deelname aan het maken van eiwitmoleculen. Met andere woorden, ze dragen informatie van de plaats waar het wordt opgeslagen in de implementatie ervan. Dienovereenkomstig duiden deze RNA's aan als informatie (IRNA) of matrix (mRNA).
Er wordt gevonden dat de IRNN-keten is gesynthetiseerd, direct met behulp van de juiste DNA-sectie als een matrix. Het gesynthetiseerde mRNA-circuit kopieert tegelijkertijd precies een van de twee DNA-circuits door zijn nucleotidesequentie (het nemen van dat de Uracil (Y) in RNA overeenkomt met zijn timinederivaat (t) in DNA). Dit gebeurt op basis van hetzelfde structurele principe van complementariteit, dat de vermindering van DNA bepaalt (Fig. 18). Het bleek dat wanneer de synthese van mRNA op DNA in de cel optreedt, dan wordt slechts één DNA-keten gebruikt als een matrix voor de vorming van een mRNA-circuit. Vervolgens komt elk G van dit DNA-circuit overeen met C in de RNA-keten in aanbouw, elke C-ketting van het DNA-circuit in het RNA-circuit, elke T-ketting - A in het RNA-circuit en elk een DNA-circuit in het RNA-circuit. Dientengevolge zal de verkregen RNA-keten strikt complementair zijn aan de DNA-matrixketen en is daarom identiek aan de nucleotidesequentie (het nemen van t \u003d y) de tweede DNA-keten. Dus de "herschrijven" van informatie van DNA op RNA vindt plaats, d.w.z. transcriptie. De combinaties "herschreven" van het nucleotide van de RNA-keten worden al rechtstreeks bepaald door de opstelling van de overeenkomstige, gecodeerde aminozuren in de eiwitketen.
Hier, zoals in de vergoeding van het gereduceerde DNA, als een van de belangrijkste momenten van het transcriptieproces, is het noodzakelijk om zijn enzymatische aard aan te geven. DNA, dat een matrix in dit proces is, bepaalt de volledige nucleotidelocatie in de synthetiserende keten van mRNA, alle specificiteit van het gevormde RNA, maar het proces zelf wordt uitgevoerd door een speciaal eiwit - het enzym. Dit enzym wordt RNA-polymerase genoemd. Het molecuul heeft een complexe organisatie waarmee het actief langs het DNA-molecuul kan bewegen, met het synthetiseren van de RNA-keten die complementair is aan een van de DNA-ketens. Het DNA-molecuul, dat dient als matrix, geeft niet uit en verandert niet, blijft in dezelfde vorm en is altijd klaar voor dergelijke herschrijven van het een onbeperkt aantal "kopieën" - mRNA. De stroom van deze MRNA's van DNA naar ribosomen is de informatiestroom die ervoor zorgt dat de programmering van de cellen van de cel van de cel, de gehele totaliteit van zijn ribo's.
Aldus beschrijft het beschouwde deel van de regeling de stroom van informatie die voortkomt uit DNA in de vorm van mRNA-moleculen tot intracellulaire deeltjes die eiwitten synthetiseren. Nu draaien we naar de stream van andere soort - naar de stroom van het materiaal waaruit het eiwit moet worden gecreëerd. Elementaire eenheden - Monomeren - eiwitmolecuul zijn aminozuren, die ongeveer 20 zijn genummerd. Om een \u200b\u200beiwitmolecuul te creëren, moeten vrije aminozuren die in de cel aanwezig zijn, betrokken zijn bij de juiste stroom die het whiteoxitheating-deeltjes binnenkomt, en er zijn al in een keten geplaatst met een zekere unieke manier daar., Dicteergegeven informatie RNA. Een dergelijke betrokkenheid van aminozuren - een bouwmateriaal voor het creëren van eiwit wordt uitgevoerd door het bevestigen van vrije aminozuren aan speciale RNA-moleculen relatief klein. Deze RNA's die dienen om gratis aminozuren aan hen te bevestigen zonder informatief te zijn, dragen een andere adapter - functie, waarvan de betekenis verder zichtbaar zal zijn. Aminozuren worden verbonden door een van de uiteinden van kleine ketens van transfers RNA (TRNA), één aminozuur per molecuul van RNA. Voor elk aminozuur in de cel zijn er zijn specifieke adapter RNA-moleculen die alleen deze aminozuren bevestigen. In een dergelijke gemonteerde vorm van aminozuren op RNA en ingevoerd in whitoxitheating-deeltjes.
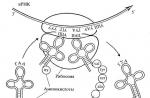
Het centrale moment van het eiwitbiosyntheseproces is de fusie van deze twee intracellulaire streams - de informatiestroom en de materiaalstroom - in de cellen van de cellen. Deze deeltjes worden ribosomen genoemd. Ribo's zijn ultramicroscopische biochemische "machines" van moleculaire maten, waarbij specifieke eiwitten worden verzameld uit de inkomende aminozuurresiduen, volgens het plan dat is gesloten in informatie-RNA. Hoewel in FIG. 19 toont slechts één deeltje, elke cel houdt duizenden ribben tegen. De hoeveelheid ribosomen bepaalt de algemene intensiteit van eiwitsynthese in de cel. De diameter van één ribosomaal deeltje is ongeveer 20 nm. Door zijn chemische aard van het ribosoma - ribonucleoproteïde: het bestaat uit een speciaal ribosomaal RNA (dit is de derde RNA-klasse die bij ons bekend is, naast informatie en adapter RNA) en moleculen van structureel ribosomaal eiwit. Samen vormt deze combinatie van verschillende dozijn macromoleculen een ideale georganiseerde en betrouwbare "auto", die een eigenschap heeft om informatie die in de mRNA-keten is gesloten en om het te realiseren in de vorm van een afgewerkt eiwitmolecuul van een specifieke structuur. Aangezien het schepsel van de werkwijze is dat lineaire opstelling van 20 verschillende aminozuren in de eiwitketen ondubbelzinnig wordt bepaald door de locatie van vier verschillende nucleotiden in de keten van het chemisch geheel ander polymeer - nucleïnezuur (mRNA), deze werkwijze voorkomt in de Ribosome is gemaakt om de term "uitzending" of "vertaling" aan te wijzen - een vertaling van het vierletterige alfabet van nucleïnezuurketens in een alfabet van een twintig gevouwen alfabet van eiwit (polypeptide) kettingen. Zoals te zien zijn, zijn alle drie bekende RNA-klassen betrokken bij het uitzendproces: een informatie-RNA, dat een uitzendobject is; Ribosomaal RNA, dat de rol speelt van een organisator van een anti-industriële ribonucleotoïde deeltje - ribosomen; En adapter RNA die de vertaler-functie uitvoeren.
|
Fig. 19. Schema van het functioneren van ribosoom |

Het eiwitsynthese-proces begint met de vorming van aminozuurverbindingen met adapter RNA-moleculen of TRNA. Tegelijkertijd gebeurde de energie "activering" van aminozuren eerst als gevolg van zijn enzymatische reactie met het adenosine-trifosfaatmolecuul (ATP), en dan is het "geactiveerde" aminozuur verbonden met het einde van een relatief kortlevende TRNA Keten, de toename van de chemische energie van het geactiveerde aminozuur is intensiteit in de vorm van de chemische bindingsenergie tussen het aminozuur en TRNA.
Tegelijkertijd is de tweede taak opgelost. Het feit is dat de reactie tussen het aminozuur en het TRA-molecuul wordt uitgevoerd door een enzym dat is aangegeven als aminoacil-tallythetase. Voor elk van de 20 aminozuren zijn er speciale enzymen die reageren met de deelname van alleen dit aminozuur. Er zijn dus ten minste 20 enzymen (aminoacyl-tall synthetase), die elk specifiek zijn voor één specifiek aminozuur. Elk van deze enzymen kan een reactie hebben die niet met elk TRNA-molecuul is, maar alleen met degenen die een strikt gedefinieerde combinatie van nucleotiden in hun keten dragen. Aldus, vanwege het bestaan \u200b\u200bvan een reeks van dergelijke specifieke enzymen die, enerzijds, de aard van het aminozuur en, aan de andere kant - de nucleotidesequentie van TRNA, blijkt elk van de 20 aminozuren te zijn " toegewezen "alleen aan een bepaald TRNA met een gegeven karakteristieke nucleotide-combinatie.
Sommige momenten van het eiwitbiosyntheseproces, voor zover wij ze vandaag presenteren, worden gegeven in FIG. 19. Hier, allereerst, het kan worden gezien dat het molecuul van een informatie-RNA is verbonden met het ribosoom of, zoals ze zeggen, het ribosoom is "geprogrammeerd" van informatie-RNA. Op het gegeven moment is direct in het ribosoom zelf slechts een relatief kort segment van de mRNA-keten. Maar het is dit segment met de deelname van ribosomen kan communiceren met adapter RNA-moleculen. En hier weer wordt de hoofdrol gespeeld door het principe van complementatie.
Dit is de verklaring van het mechanisme van de reden waarom deze tript van de mRNA-keten overeenkomt met een strikt gedefinieerd aminozuur. De nodige tussenliggende link of adapter, met de "erkenning" van elk aminozuur van zijn triplet op mRNA is Adapter RNA (TRNA).
In FIG. 19 Het kan worden gezien dat in het ribosoom, in aanvulling op het TRA-molecuul met een gehangen aminozuur, er een ander TRA-molecuul is. Maar in tegenstelling tot het hierboven besproken TRNA-molecuul, is dit molecuul-tRNA aan het einde bevestigd aan het einde van de synthese van eiwit (polypeptide) keten. Deze bepaling weerspiegelt de dynamiek van gebeurtenissen die voorkomen in het ribosoom tijdens de synthese van het eiwitmolecuul. Deze dynamiek kan als volgt worden ingebeeld. Laten we beginnen met een bepaald tussenliggende koppel dat wordt weerspiegeld in FIG. 19 en gekenmerkt door de aanwezigheid van een reeds begonnen te worden gebouwd door een eiwitketen die aan haar TRNA is bevestigd en net in riboSoom is aangegaan en verbonden is met een triplet van een nieuw TRA-molecuul met het overeenkomstige aminozuur. Blijkbaar leidt de handeling van bevestiging van het TRNA-molecuul aan het ribosoom-gelegen ribosoom-triplet tot een dergelijke wederzijdse oriëntatie en nauw contact tussen het aminozuurresidu en het eiwitcircuit in aanbouw, dat een covalente band tussen hen voorkomt. Communicatie ontstaat op een zodanige manier dat het einde van de eiwitketen in aanbouw (in Fig. 19 aan TRNA) wordt overgedragen van deze TRNA naar het aminozuurresidu van de Aminoacyl-handel. Als gevolg van het "juiste" TRNA is het spelen van de rol van de "donor", vrij en de eiwitketen - overgedragen aan de "acceptor", d.w.z. Op de "linker" (ontvangen) aminoacil-TRNA. Dientengevolge zal de eiwitketen worden langwerpig per aminozuur en bevestigd aan de "linker" TRNA. Hierna wordt het "linker" TRNA overgedragen samen met het mRNA-nucleotide-gerelateerde triplet naar rechts, dan wordt het voormalige "donor" -molecuul van hier voorzien van hier en laat ribosomen achter. In haar plaats verschijnt een nieuw TRNA met een proteïne keten in aanbouw die is uitgebreid tot één aminozuurresidu, en de mRNA-keten zal ten opzichte van het ribosoom op één triplet rechts worden bevorderd. Als gevolg van de bevordering van de MRNA-keten verschijnt één triplet naar rechts in het ribosoom de volgende vacante triplet (uou) en de bijbehorende TRNA met aminozuur (fenylalanyl-tRNA) zal er onmiddellijk aan worden toegevoegd volgens de Aanvullend principe. Dit zal opnieuw veroorzaken dat de vorming van een covalente (peptide) verbinding tussen de eiwitketen in aanbouw en het fenylalanine-residu en volgde de promotie van de mRNA-keten tot één triplet rechts met alle gevolgen van hier, enz. Op deze manier wordt het consequent uitgevoerd, een triplet voor een triplet, waarbij de keten van een informatie-RNA door het ribosoom wordt getrokken, waardoor de Irnn-keten door het ribosoom volledig is, van begin tot einde. Tegelijkertijd is er een consistent aminozuur voor het aminozuur, de eiwitketen conjugy. Dienovereenkomstig komt in het ribosoma de een na de ander de moleculen van de TRNA met aminozuren en komen de TRA-moleculen uit zonder aminozuren. Dekking in de oplossing buiten de ribosomen, de vrije TRA-moleculen zijn weer verbonden met aminozuren en dragen ze opnieuw in ribosoma, waardoor cyclisch wordt verwijzend zonder vernietiging en verandering.
Het belangrijkste postulaat van moleculaire biologie
Er zijn drie moleculaire biologieprocessen.
Verworven formulier
Deze vaker vorm van orotatacidurië kan worden waargenomen:
· Met elk defect enzymen van ureumsynthesebehalve carbamoyl fosfaatynthetase. Tegelijkertijd komt mitochondriale carbamoylfosfaat (in de norm die wordt gebruikt voor de vorming van ureum) uit hen en wordt het gebruikt voor overmatige of gemaakte zure synthese. De ziekte is meestal vergezeld van bygipermonmoniamia,
· Bij de behandeling van alopurinol jicht, die kan worden in een oxyfarinolmonucleotide, die een orotatdecarboxylase-remmer is, die opnieuw leidt tot de accumulatie van orotata.
De belangrijkste figuur van matrixbiosynthese is nucleïnezuur-RNA en DNA. Het zijn polymeermoleculen die stikstofhoudende basen van vijf typen, pentoses van twee soorten en residuen van fosforzuur omvatten. Stikstofbases in nucleïnezuren kunnen purin zijn ( adenin,guaniaans) en pyrimidine ( cytozin, uracil (alleen in RNA) timin (alleen in DNA)). Afhankelijk van de koolhydraatstructuur, markeer dan ribonucleïnezuren - Bevatten Ribosa (RNA), en deoxyribonucleïnezuren - Bevatten deoxyribose (DNA).
Term " matrix biosynthese"Het impliceert het vermogen van de cel om polymeermoleculen te synthetiseren, zoals nucleïnezuren en eiwitten, op basis van sjabloon - matrians. Dit zorgt voor een nauwkeurige overdracht van de meest complexe structuur uit de reeds bestaande moleculen naar het nieuw inbegrepen.
In de overweldigende meerderheid van de gevallen wordt de overdracht van erfelijke informatie uit de moedercel naar een dochteronderneming uitgevoerd met behulp van DNA ( replicatie). Om genetische informatie te gebruiken, vereist de cel zelf RNA gevormd op de DNA-matrix ( transcriptie). Verder is RNA direct betrokken bij alle stadia van de synthese van eiwitmoleculen ( uitzending), het verschaffen van de structuur en activiteit van de cel.
Op het voorgaande is gebaseerd centrale Dogma Molecular BiologyVolgens welke de overdracht van genetische informatie alleen wordt uitgevoerd op nucleïnezuur (DNA en RNA). De ontvanger van de informatie kan ander nucleïnezuur (DNA of RNA) en eiwit zijn.
Hybridisatie is al veel gebruikt
Als u de DNA-oplossing boven de temperatuur van 90 ° C verwarmt of de pH wordt verschoven in een sterk alkalisch of scherp zijden, worden de waterstofbindingen tussen de DNA-draden vernietigd en is de dubbele helix gebroken. Gebeurt dNA-denaturatieof, op een andere manier, smeltend. Als u een agressieve factor verwijdert, gebeurt dan renaturaof uitgloeien. Tijdens het gloeiende draden "Vind" complementaire gebieden van elkaar en verschijnen uiteindelijk in een dubbele helix.
Als in een "buis" smelten en gloeien van het DNA-mengsel, bijvoorbeeld, mensen muisSommige delen van de Mouse DNA-circuits worden herenigd met complementaire gebieden van menselijke DNA-ketens met onderwijs hybrida. Het aantal van dergelijke sites is afhankelijk van de mate van verwantschap van soorten. Hoe dichter de standpunten tussen zichzelf, hoe groter de complementariteit van de DNA-draden. Dit fenomeen wordt genoemd dNA DNA-hybridisatie.
Als RNA in de oplossing aanwezig is, kunt u implementeren dNA RNA-hybridisatie. Dergelijke hybridisatie helpt bij het vaststellen van de nabijheid van bepaalde DNA-sequenties met een RNA.
Hybridisatie van DNA-DNA en DNA-RNA wordt gebruikt als effectieve tool in moleculaire genetica, forensische geneeskunde, antropologie om genetische verwantschap tussen soorten vast te stellen.
Cel als zodanig bezit een enorm aantal verschillende functies, zoals we al hebben gesproken, sommige van hen zijn over het algemeen cel, sommige zijn speciaal, kenmerkend voor speciale celtypen. De belangrijkste werkmechanismen voor het uitvoeren van deze functies zijn eiwitten of hun complexen met andere biologische macromoleculen, zoals nucleïnezuren, lipiden en polysacchariden. Het is dus bekend dat de transportprocessen in verschillende stoffen, beginnend met ionen, eindigend met macromoleculen, worden bepaald door de werking van speciale eiwitten of lipoproteïnecomplexen in de samenstelling van plasma en andere celmembranen. Vrijwel alle processen van synthese, verval, herstructurering van verschillende eiwitten, nucleïnezuren, lipiden, treedt koolhydraten op als gevolg van activiteit die specifiek is voor elke individuele reactie van enzymeiwitten. Synthese van individuele biologische monomeren, nucleotiden, aminozuren, vetzuren, suikers, enz. Worden ook uitgevoerd door een enorm aantal specifieke enzymen - eiwitten. De vermindering die leidt tot celmobiliteit of aan de beweging van stoffen en structuren in de cellen wordt ook uitgevoerd door speciale contractiele eiwitten. Veel celreacties in reactie op de impact van externe factoren (virussen, hormonen, buitenaardse eiwitten, enz.) Begint met de interactie van deze factoren met speciale cel-eiwittenreceptoren.
Eiwitten zijn de belangrijkste componenten van bijna alle cellulaire structuren. Veel chemische reacties in de cel worden bepaald door de set enzymen, die elk een of meer individuele reacties leiden. De structuur van elk individueel eiwit wordt strikt gespecificeerd, dat wordt uitgedrukt in de specificiteit van hun primaire structuur - in de aminozuursequentie langs het polypeptide, eiwitketen. Bovendien is de specificiteit van deze aminozuursequentie onmiskenbaar herhaald in alle moleculen van dit cellulaire eiwit.
Een dergelijke correctheid bij het reproduceren van de ondubbelzinnige sequentie van aminozuren in de eiwitketen wordt bepaald door de structuur van het DNA van het gengedeelte, dat uiteindelijk verantwoordelijk is voor de structuur en synthese van dit eiwit. Deze ideeën dienen als de belangrijkste postulate van moleculaire biologie, het "dogma". Informatie over het toekomstige eiwitmolecuul wordt verzonden naar de locatie van zijn synthese (in ribosomen) door een tussenpersoon - informatie-RNA (IRNA), waarvan de nucleotidesamenstelling de samenstelling en volgorde van het nucleotide van het DNA-gen weerspiegelt. Een polypeptideketen is gebouwd in het ribosoom, de sequentie van aminozuren waarin wordt bepaald door de sequentie van nucleotiden in de IRNA, de reeks van hun triplets. Aldus benadrukt het centrale dogma van de moleculaire biologie de unidirectionaliteit van informatieoverdracht: alleen van DNA tot eiwit, met behulp van intermediaire, IRNA (DNA ® IRNK ®-eiwit). Voor sommige RNA-bevattende virussen kan het informatietransmissiekring volgens het RNA-schema gaan - Irnk - eiwit. Het verandert de essentie van de zaak niet, omdat de bepalende link hier ook nucleïnezuur is. Omgekeerde manieren van vastberadenheid van eiwit tot nucleïnezuur, naar DNA of RNA zijn onbekend.
Om verder te gaan naar de studie van celstructuren die geassocieerd zijn met alle stadia van eiwitsynthese, moeten we in het kort verblijven op de belangrijkste processen en componenten die dit fenomeen bepalen.
Momenteel kan op basis van moderne ideeën over de biosynthese van eiwitten, de volgende algemene hoofdschema van dit complexe en meertrapsproces worden gegeven (fig. 16).
Het belangrijkste, "team", de rol bij het bepalen van de specifieke structuur van eiwitten behoort tot deoxyribonucleïnezuur - DNA. Het DNA-molecuul is een extreem lange lineaire structuur bestaande uit twee intercloine-polymeerketens. Samengestelde elementen - Monomeren - deze kettingen zijn vier variëteiten van deoxyribonucleotiden, een afwisseling of een reeks waarvan langs de keten uniek en specifiek is voor elk DNA-molecuul en elk van zijn secties. Verschillende voldoende lange delen van DNA-moleculen zijn verantwoordelijk voor de synthese van verschillende eiwitten. Een DNA-molecuul kan dus de synthese bepalen van een groot aantal functioneel en chemisch verschillende cel-eiwitten. Voor de synthese van elk type eiwitten is slechts een bepaald deel van het DNA-molecuul verantwoordelijk. Een dergelijk deel van het DNA-molecuul dat is geassocieerd met de synthese van één eiwit in de cel wordt vaak aangeduid met de term "CYSTRON". Momenteel wordt het concept van Cyastron beschouwd als een equivalent concept van gen. In de unieke genstructuur, in een bepaalde sequentiële opstelling van zijn nucleotiden langs de keten, wordt alle informatie over de structuur van één overeenkomstig eiwit afgesloten.
Uit de algemene regeling van eiwitsynthese wordt het gezien (zie figuur 16) dat het initiële item waaruit de informatiestroom voor de biosynthese van eiwitten in de cel begint, DNA is. Daarom is het het DNA dat de eerste verslag van informatie bevat die moet worden gehandhaafd en gereproduceerd van de cel naar de cel, van generatie tot generatie.
Kort aanraken van de vraag over de plaats van opslag van genetische informatie, d.w.z. Op de lokalisatie van DNA in de cel kunnen we het volgende zeggen. Het is al lang bekend dat het DNA, in tegenstelling tot alle andere componenten van het Whiteoxithating-apparaten, een speciale, zeer beperkte lokalisatie heeft: de locatie in de cellen van hogere (eukaryote) organismen zal de celkern zijn. In de lagere (prokaryote) organismen die geen ingerichte celkern hebben, wordt DNA ook opgemerkt van de rest van het protoplasma in de vorm van een of meer compacte nucleotidevormingen. In volledige naleving hiermee is de eukaryote kernel of de prokaryote nucleïde al lang beschouwd als een aanbieder van genen als een unieke cellulaire organoïde, die de implementatie van erfelijke tekenen van organismen en hun transfer tot generaties controleert.
Het basisprincipe dat ten grondslag ligt aan de macromoleculaire structuur van DNA is het zogenaamde complementaire principe (fig. 17). Zoals reeds vermeld, bestaat het DNA-molecuul uit twee interclowed-ketens. Deze ketens zijn met elkaar verbonden door de interactie van hun tegenovergestelde nucleotiden. Tegelijkertijd is volgens structurele redenen het bestaan \u200b\u200bvan een dergelijke kruik-in-kamerstructuur alleen mogelijk als de tegenovergestelde nucleotiden van beide ketens sterisch complementair zullen zijn, d.w.z. zal je ruimtelijke structuur zijn om elkaar aan te vullen. Een dergelijke complementaire - complementaire - paren nucleotiden zijn stoom A-T (Adenin-Timin) en een paar van Mr. (Guanin-Cytosin).
Daarom, volgens dit principe van complementariteit, als we een bepaalde sequentie van vier variëteiten van nucleotiden in één keten hebben, dan in de tweede keten, zal de sequentie van nucleotiden ondubbelzinnig worden bepaald, zodat elk en de eerste keten overeenkomt met de Tweede ketting, elke eerste keten - en in de tweede keten, is elke G van de eerste keten C in de tweede keten en elke C eerste keten - R in de tweede keten.
Er kan worden gezien dat het opgegeven structurele principe die ten grondslag ligt aan de schuimstructuur van het DNA-molecuul het gemakkelijk maakt om de exacte reproductie van de bronstructuur, d.w.z. te begrijpen Exacte weergave van informatie die is opgenomen in de molecuulcircuits in de vorm van een bepaalde sequentie van 4 variëteiten van nucleotiden. Inderdaad, de synthese van nieuwe DNA-moleculen in de cel gebeurt alleen op basis van de reeds bestaande DNA-moleculen. Tegelijkertijd beginnen de twee ketens van het oorspronkelijke DNA-molecuul zich te verspreiden van een van de uiteinden, en op elk van de gescheiden single-secties begint te worden verzameld uit de tweede keten die aanwezig is in het medium vrije nucleotiden bij nauwkeurig naleving met het principe van complementariteit. Het proces van discrepantie tussen twee ketens van het oorspronkelijke DNA-molecuul gaat door, en dienovereenkomstig worden beide ketens aangevuld met aanvullende circuits. Als gevolg hiervan, zoals te zien in het diagram verschijnen twee DNA-moleculen in plaats van één, precies identiek aan het origineel. In elk van de resulterende "dochteronderneming" DNA-molecuul is één ketting, zoals te zien, volledig te wijten aan het origineel, en de andere is opnieuw gesynthetiseerd.
Het belangrijkste is dat het nodig is om opnieuw te benadrukken, dit is dat het potentiële vermogen om nauwkeurig te reproduceren wordt gelegd in de kleinste complementaire DNA-structuur als zodanig, en de ontdekking hiervan is absoluut een van de belangrijkste prestaties van de biologie.
Het probleem van reproductie (reductioneel) DNA is echter niet uitgeput door de statuut van het potentiële vermogen van de structuur om zijn nucleotidesequentie nauwkeurig te reproduceren. Het feit is dat het DNA zelf helemaal geen zelfreproductiemolecuul is. Om het proces van synthese - DNA-reproductie volgens de hierboven beschreven regeling uit te voeren, is de activiteit van een speciaal enzymatisch complex dat het DNA-polymerase wordt genoemd noodzakelijk. Blijkbaar is het dit enzym dat het DNA-molecuul constant spreekt van het ene uiteinde naar een ander proces van discrepanties van twee circuits met gelijktijdige polymerisatie op hen vrij van nucleotiden door aanvullend principe. Aldus stelt DNA, zoals een matrix, alleen de volgorde van het nucleotide in de synthetiserende ketens, en het proces zelf leidt eiwit. Het werk van het enzym tijdens de vermindering van DNA is een van de meest interessante problemen vandaag. Blijkbaar lijkt het DNA-polymerase actief te kruipen langs het twintigste molecuul DNA van het ene uiteinde naar het andere, waardoor een gespleten verminderde "staart" achterblijft. De fysieke principes van dit werk van dit eiwit zijn nog niet duidelijk.
DNA en zijn individuele functionele secties die informatie over de structuur van eiwitten dragen, directe deelname zelf in het proces van het maken van eiwitmoleculen worden niet geaccepteerd. De eerste fase op weg naar de uitvoering van deze informatie die is opgenomen in DNA-kettingen is het zogenaamde transcriptieproces of "herschrijven". In dit proces is de DNA-ketens, zoals op de matrix, een synthese van een chemisch gerelateerd polymeer - ribonucleïnezuur (RNA). Het RNA-molecuul is één ketting, waarvan de monomeren vier variëteiten van ribonucleotiden zijn, die worden beschouwd als een kleine wijziging van de vier variëteiten van DNA Deoxyribonucleotiden. De sequentie van de vier variëteiten van ribonucleotiden in het resulterende RNA-circuit herhaalt exact de sequentie van de overeenkomstige deoxyribonucleotiden van een van de twee DNA-circuits. Op deze manier wordt de nucleotidesequentie van genen gekopieerd als RNA-moleculen, d.w.z. De informatie die is opgenomen in de structuur van dit gen is volledig herschreven op RNA. Een groot, theoretisch onbeperkt aantal van dergelijke "kopieën" - RNA-moleculen kan uit elk gen worden verwijderd. Deze moleculen, herschreven in vele exemplaren van beide "kopieën" van genen en dragen daarom dezelfde informatie als genen, uiteenlopen door de cel. Ze zijn al direct bij de cellen van de cellen opgenomen en accepteren "Persoonlijke" deelname aan het maken van eiwitmoleculen. Met andere woorden, ze dragen informatie van de plaats waar het wordt opgeslagen in de implementatie ervan. Dienovereenkomstig duiden deze RNA's aan als informatieve of matrix RNA's, afgekort mRNA (of IRNA).
Er blijkt dat het informatie-RNA-circuit wordt gesynthetiseerd, direct met behulp van het juiste DNA-gedeelte als matrix. Het gesynthetiseerde mRNA-circuit kopieert tegelijkertijd precies een van de twee DNA-circuits door zijn nucleotidesequentie (het nemen van dat de Uracil (Y) in RNA overeenkomt met zijn timinederivaat (t) in DNA). Dit gebeurt op basis van hetzelfde structurele principe van complementariteit, dat de vermindering van DNA bepaalt (Fig. 18). Het bleek dat wanneer de synthese van mRNA op DNA in de cel optreedt, dan wordt slechts één DNA-keten gebruikt als een matrix voor de vorming van een mRNA-circuit. Vervolgens komt elk G van dit DNA-circuit overeen met C in de RNA-keten in aanbouw, elke C-ketting van het DNA-circuit in het RNA-circuit, elke T-ketting - A in het RNA-circuit en elk een DNA-circuit in het RNA-circuit. Dientengevolge zal de resulterende RNA-keten strikt complementair zijn aan de DNA-matrixketen en daarom identiek aan de sequentie van nucleotiden (het nemen van t \u003d y) de tweede DNA-keten. Dus de "herschrijven" van informatie van DNA op RNA vindt plaats, d.w.z. transcriptie. De combinaties "herschreven" van het nucleotide van de RNA-keten worden al rechtstreeks bepaald door de opstelling van de overeenkomstige, gecodeerde aminozuren in de eiwitketen.
Hier, zoals in de vergoeding van het gereduceerde DNA, als een van de belangrijkste momenten van het transcriptieproces, is het noodzakelijk om zijn enzymatische aard aan te geven. DNA, dat een matrix in dit proces is, bepaalt de volledige nucleotidelocatie in de synthetiserende keten van mRNA, alle specificiteit van het gevormde RNA, maar het proces zelf wordt uitgevoerd door een speciaal eiwit - het enzym. Dit enzym wordt RNA-polymerase genoemd. Het molecuul heeft een complexe organisatie waarmee het actief langs het DNA-molecuul kan bewegen, met het synthetiseren van de RNA-keten die complementair is aan een van de DNA-ketens. Het DNA-molecuul, dat dient als matrix, geeft niet uit en verandert niet, blijft in dezelfde vorm en is altijd klaar voor zo'n herschrijven van het een onbeperkt aantal "kopieën" - mRNA. De stroom van deze MRNA's van DNA naar ribosomen is de informatiestroom die ervoor zorgt dat de programmering van de cellen van de cel van de cel, de gehele totaliteit van zijn ribo's.
Aldus beschrijft het beschouwde deel van de regeling de stroom van informatie die voortkomt uit DNA in de vorm van mRNA-moleculen tot intracellulaire deeltjes die eiwitten synthetiseren. Nu draaien we naar de stream van andere soort - naar de stroom van het materiaal waaruit het eiwit moet worden gecreëerd. Elementaire eenheden - Monomeren - eiwitmolecuul zijn aminozuren met 20 verschillende variëteiten. Om een \u200b\u200beiwitmolecuul (synthetisering) te creëren, moeten gratis aminozuren die in de cel aanwezig zijn, betrokken zijn bij de juiste stroom die het whiteoxitheating-deeltjes binnenkomen, en er zijn al in een keten geplaatst met een bepaalde unieke manier die wordt gedicteerd door informatie-RNA. Deze betrokkenheid van aminozuren - een bouwmateriaal voor het creëren van eiwit wordt uitgevoerd door de toevoeging van vrije aminozuren aan speciale RNA-moleculen relatief klein. Deze RNA's die dienen om deze gratis aminozuren aan te sluiten, zijn niet informerend en dragen een andere adapter - functie, waarvan de betekenis verder zichtbaar zal zijn. Aminozuren worden verbonden door een van de uiteinden van kleine ketens van transfers RNA (TRNA), één aminozuur per molecuul van RNA.
Voor elke verscheidenheid aan aminozuren in de cel zijn er zijn specifiek, die alleen deze variëteit aan aminozuren van het adapter RNA-molecuul bevestigt. In een dergelijke vorm bezocht op RNA, aminozuren en betreed de witteogige deeltjes.
Het centrale moment van het eiwitbiosyntheseproces is de fusie van deze twee intracellulaire streams - de informatiestroom en de materiaalstroom - in de cellen van de cellen. Deze deeltjes worden ribosomen genoemd. Ribo's zijn ultramicroscopische biochemische "machines" van moleculaire maten, waarbij specifieke eiwitten worden verzameld uit de inkomende aminozuurresiduen, volgens het plan dat is gesloten in informatie-RNA. Hoewel in deze regeling (fig. 19) slechts één deeltje toont, houdt elke cel duizenden ribben tegen. De hoeveelheid ribosomen bepaalt de algemene intensiteit van eiwitsynthese in de cel. De diameter van één ribosomaal deeltje is ongeveer 20 nm. Door zijn chemische aard van het ribosoma - ribonucleoproteïde: het bestaat uit een speciaal ribosomaal RNA (dit is de derde RNA-klasse die bij ons bekend is, naast informatie en adapter RNA) en moleculen van structureel ribosomaal eiwit. Samen vormt deze combinatie van verschillende dozijn macromoleculen een ideale georganiseerde en betrouwbare "auto", die een eigenschap heeft om informatie die in de mRNA-keten is gesloten en om het te realiseren in de vorm van een afgewerkt eiwitmolecuul van een specifieke structuur. Aangezien het schepsel van de werkwijze is dat de lineaire opstelling van 20 variëteiten van aminozuren in de eiwitketen uniek wordt bepaald door de locatie van de vier variëteiten van nucleotiden in het chemisch geheel ander polymeerpolymeer (mRNA), dan is dit proces in de Ribosome is gemaakt om de term "uitzending" of "vertaling" aan te wijzen - een vertaling van een 1-letterig alfabet van nucleïnezuurketens door een alfabet van 20-letterige eiwit (polypeptide) kettingen. Zoals te zien zijn, zijn alle drie bekende RNA-klassen betrokken bij het uitzendproces: een informatieve RNA, dat een vertaalobject, ribosomaal RNA is, dat de rol speelt van een organisator van een anti-ijverse ribonucleotopoïde deeltje - ribosoom en adapter RNA het uitvoeren van de vertaler-functie.
Het eiwitsynthese-proces begint met de vorming van aminozuurverbindingen met adapter RNA-moleculen of TRNA. Tegelijkertijd gebeurde de energie "activering" van aminozuren eerst als gevolg van zijn enzymatische reactie met het adenosine-trifosfaatmolecuul (ATP), en dan is het "geactiveerde" aminozuur verbonden met het einde van een relatief kortlevende TRNA Keten, de toename van de chemische energie van het geactiveerde aminozuur is intensiteit in de vorm van de chemische bindingsenergie tussen het aminozuur en TRNA.
Maar tegelijkertijd is de tweede taak opgelost. Het feit is dat de reactie tussen het aminozuur en het TRA-molecuul wordt uitgevoerd door een enzym dat is aangegeven als aminoacil-tallythetase. Voor elk van de 20 variëteiten van aminozuren zijn er speciale enzymen die reageren met de deelname van alleen dit aminozuur. Er zijn dus ten minste 20 enzymen (aminoacyl-trnk synthetase), die elk specifiek zijn voor een verscheidenheid aan aminozuren. Elk van deze enzymen kan een reactie hebben die niet met elk TRNA-molecuul is, maar alleen met degenen die een strikt gedefinieerde combinatie van nucleotiden in hun keten dragen. Dus, vanwege het bestaan \u200b\u200bvan een reeks van dergelijke specifieke enzymen die, enerzijds, de aard van het aminozuur en, aan de andere kant - de nucleotidesequentie van TRNA, blijkt elk van de 20 variëteiten van aminozuren "Toegewezen" alleen aan een bepaald TRNA met een gegeven karakteristieke nucleotide-combinatie.
Sommige momenten van het eiwitbiosyntheseproces, voor zover wij ze vandaag presenteren, worden gegeven in FIG. negentien.
Hier, allereerst, het is te zien dat het informatie-RNA-molecuul is verbonden met het ribosoom of, zoals ze zeggen, het ribosoom "geprogrammeerd" -informatie-RNA. Op een bepaald moment is direct in het ribosoom zelf slechts een relatief kort segment van de mRNA-keten. Maar het is dit segment met de deelname van ribosomen kan communiceren met adapter RNA-moleculen. En hier wordt de hoofdrol tweemaal het bovengenoemde principe van complementatie gespeeld.
Dit is de verklaring van het mechanisme van de reden waarom deze tript van de mRNA-keten overeenkomt met een strikt gedefinieerd aminozuur. Het is te zien dat de noodzakelijke tussenliggende link, of de adapter, met de "erkenning" van elk aminozuur van zijn triplet op mRNA-adapter RNA (TRNA) is.
Verder in het diagram (zie fig. 19) is het gezien dat in het ribosoom, naast de bovenstaande-beschouwde TRNA-moleculen met een gehangen aminozuur, er een ander TRA-molecuul is. Maar in tegenstelling tot het hierboven besproken TRNA-molecuul, wordt dit molecuultRNA bevestigd aan het einde van het eiwit (polypeptide) keten in het proces van synthese. Deze bepaling weerspiegelt de dynamiek van gebeurtenissen die voorkomen in het ribosoom tijdens de synthese van het eiwitmolecuul. Deze dynamiek kan als volgt worden ingebeeld. Laten we beginnen met een bepaald tussenliggende koppel dat in het diagram wordt weerspiegeld en gekenmerkt door de aanwezigheid van een eiwitketen die al is gebouwd, gehecht aan haar TRNA en zojuist in ribosoma is ingeschakeld en verbonden is met een triplet van een nieuw TRNA-molecuul met de overeenkomend aminozuur. Blijkbaar leidt de handeling van bevestiging van het TRNA-molecuul aan het ribosoom-gelegen ribosoom-triplet tot een dergelijke wederzijdse oriëntatie en nauw contact tussen het aminozuurresidu en het eiwitcircuit in aanbouw, dat een covalente band tussen hen voorkomt. Communicatie treedt op zodanig gebeurd dat het einde van de eiwitketen in aanbouw, op het diagram dat aan de TRNA is bevestigd, wordt overgedragen van deze TRNA naar het aminozuurrest van de toegelaten Aminoacil-TRNA. Als gevolg van het "juiste" TRNA zal het spelen van de rol van de "donor", vrij zijn en de eiwitketen - een "linker" (ontvangen) aminoacyl-tRNA - "links" (ontvangen) tot "acceptor" - Dientengevolge zal de eiwitketen worden verlengd per aminozuur en bevestigd aan de "links» TRNA. Hierna wordt het "links" TRNA aan het overbrengen samen met het mRNA-rechter nucleotide-gerelateerde triplet, dan wordt het voormalige "donor" -molecuul van hier verstoken en laat het ribosoma, een nieuwe TRNA met een eiwit dat in aanbouw aminozuurresidu wordt geketend , en de mRNA-keten zal worden gepromoveerd ten opzichte van het ribosoom op één triplet rechts. Als gevolg van de bevordering van de MRNA-keten verschijnt één triplet naar rechts in het ribosoom de volgende vacante triplet (uou) en de bijbehorende TRNA met aminozuur (fenylalanyl-tRNA) zal er onmiddellijk aan worden toegevoegd volgens de Aanvullend principe. Dit zal opnieuw veroorzaken dat de vorming van een covalente (peptide) verbinding tussen de eiwitketen in aanbouw en het fenylalanine-residu en volgde de promotie van de mRNA-keten tot één triplet rechts met alle gevolgen van hier, enz. Op deze manier wordt het consequent uitgevoerd, een triplet voor een triplet, waarbij de keten van een informatie-RNA door ribosoom wordt getrokken, waardoor de Irnn-keten "Ribosome volledig, van begin tot eind wordt gelezen. Tegelijkertijd is er een consistent aminozuur voor het aminozuur, de eiwitketen conjugy. Dienovereenkomstig komt in het ribosoma de een na de ander de moleculen van de TRNA met aminozuren en komen de TRA-moleculen uit zonder aminozuren. Dekking in de oplossing buiten de ribosomen, de vrije TRA-moleculen zijn weer verbonden met aminozuren en dragen ze opnieuw in ribosoma, waardoor cyclisch wordt verwijzend zonder vernietiging en verandering.
De belangrijkste figuur van matrix biosynthese is rNA en DNA-nucleïnezuren . Het zijn polymeermoleculen die stikstofhoudende basen van vijf typen, pentoses van twee soorten en residuen van fosforzuur omvatten. Stikstofbases in nucleïnezuren kunnen purin (adenine, guanine) en pyrimidine (cytosine, uracil (alleen in RNA), Timin (alleen in DNA)) zijn. Afhankelijk van de koolhydraatstructuur, markeer dan ribonucleïnezuren - Bevatten Ribosa (RNA), en deoxyribonucleïnezuren - Bevatten deoxyribose (DNA).
Term " matrix biosynthese"Het impliceert het vermogen van de cel om polymeermoleculen te synthetiseren, zoals nucleïnezuren en eiwitten, op basis van sjabloon - matrix. Dit zorgt voor een nauwkeurige overdracht van de meest complexe structuur uit de reeds bestaande moleculen naar het nieuw inbegrepen.
Het belangrijkste postulaat van moleculaire biologie
In de overweldigende meerderheid van de gevallen wordt de overdracht van erfelijke informatie van de maternalecel naar een dochteronderneming uitgevoerd met behulp van DNA (replicatie). Om genetische informatie te gebruiken, vereist de cel zelf RNA gevormd op de DNA-matrix (transcriptie). Verder is RNA direct betrokken bij alle stadia van de synthese van eiwitmoleculen (vertaling), die de structuur en activiteit van de cel verschaft.
Op het voorgaande is gebaseerd centrale Dogma Molecular BiologyVolgens welke de overdracht van genetische informatie alleen wordt uitgevoerd op nucleïnezuur (DNA en RNA). De ontvanger van de informatie kan ander nucleïnezuur (DNA of RNA) en eiwit zijn.